Aktivní transport
V buněčné biologii se aktivním transportem označuje pohyb molekul přes buněčné membrány z oblasti nižší koncentrace do oblasti vyšší koncentrace těchto molekul, tedy proti koncentračnímu gradientu. Aktivní transport proto vyžaduje spotřebu buněčné energie. Rozlišují se dva typy aktivního transportu: primární aktivní transport, který využívá energii v podobě ATP, a sekundární aktivní transport, který využívá elektrochemický protonový gradient. Příkladem aktivního transportu je např. vstřebávání glukosy v tenkém střevě nebo minerálů do kořenových vlásků rostlin.[1]
Historie
Už v roce 1848 navrhl německý fyziolog Emil du Bois-Reymond možnost aktivního transportu látek skrz membrány.[2] V roce 1948 formuloval Thomas Rosenberg koncept aktivního transportu na základě energetických bilancí, který ale později ještě přeformuloval.[3]
V roce 1957 objevil dánský vědec Jens Christian Skou důležitý transmembránový enzym Na+/K+-ATPázu, za jejíž studium získal v roce 1997 Nobelovu cenu za chemii.[4]
Významný byl také objev symportu sodík-glukosa roku 1960 americkým vědcem Robertem K. Cranem. Tento transport probíhá zejména v tenkém střevě a zkoumá se hlavně ve spojitosti s léčbou diabetu.[5]
Mechanismus
Specializované transmembránové proteiny mají receptory, které rozeznávají a navážou danou molekulu nebo iont a umožní jejich průchod skrz membránu. Za normálních okolností je membrána pro látky neprůchodná buď kvůli fosfolipidové dvojvrstvě, nebo kvůli opačnému koncentračnímu gradientu dané látky. Primární aktivní transport zajišťují proteiny (tzv. pumpy), které využívají energii ve formě ATP. Proteiny sekundárního aktivního transportu využívají potenciální energii vzniklou využitím elektrochemického gradientu. Energie vzniklá pohybem jedné látky směrem "dolů" svým koncentračním gradientem je využita pro transport jiné látky proti svému koncentračnímu gradientu. To je rozdíl oproti pasivnímu transportu, který nepotřebuje buněčnou energii a přenos látek jede vždy ve směru jejich koncentračního gradientu.[6]
Primární aktivní transport
Primárním aktivním transportem jsou přenášeny hlavně ionty H+, Na+, K+, Mg2+ a Ca2+, které k průchodu membránou potřebují iontové pumpy nebo iontové kanály. Většinou jsou to transmembránové enzymy ATPázy. Tou nejdůležitější je sodno-draselná pumpa (Na+/K+-ATPáza), která pomáhá udržovat buněčný potenciál. Tato pumpa zajišťuje přenos tří Na+ iontů ven z buňky výměnou za dva K+ ionty dovnitř buňky za spotřeby ATP.[7]
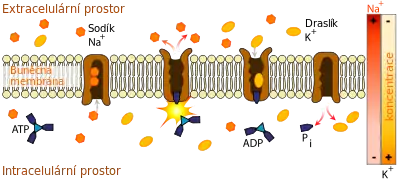
ATP se nejprve hydrolyzuje na ADP a fosfátový zbytek. Fosfát se naváže na transportní protein (fosforyluje ho) a zároveň se naváže přenášený iont, což způsobí konformační změnu transportního proteinu a dojde k přenosu iontu skrz membránu. Nakonec se hydrolyzuje navázaný fosfát a transportní protein se vrátí do původního stavu.[8]
Dalšími zdroji energie pro primární aktivní transport je redoxní energie a světelná energie. Redoxní energii využívá elektronový transportní řetězec v mitochondriích, kde se redukční energie molekul NADH využívá k pohybu protonů H+ do mezimembránového prostoru proti jejich koncentračnímu gradientu. Světelná energie se využívá při fotosyntéze k vytvoření protonového gradientu skrz membránu tylakoidů a zároveň k vytvoření redukční energie ve formě NADPH.[9]
Typy ATPáz
ATPázy jsou transmembránové enzymy, které hydrolyzují ATP na ADP a fosfátový zbytek. Energii vzniklou tímto štěpením využívají ke své činnosti.
- P-ATPázy
Tento typ ATPáz dostal označení P podle schopnosti fosforylace části vlastní struktury obsahující aspartát. Příkladem jsou protonové pumpy (H+ -ATPázy) v rostlinných buňkách, vodno-draselná pumpa (H+/K+ -ATPáza) v žaludku pro udržení kyselého pH, nebo Ca2+-ATPáza SERCA ve svalových buňkách umožňující relaxaci svalu.
Nejvýznamnější je již zmíněná sodno-draselná pumpa (Na+/K+-ATPáza), kdy jsou 3 ionty Na+ vyměněny za 2 ionty K+. Oba ionty se pohybují proti svému koncentračnímu gradientu a v buňce vzniká záporný náboj. Tím se také udržuje tzv. klidový membránový potenciál.[10]
Tzv. vakuolární ATPázy se vyskytují např. v endozomech, lysozomech, vakuolách, nebo např. osteoklastech. Jejich funkcí je hlavně udržování pH.
- F-ATPázy
Jsou to především ATP syntázy, tedy enzymy využívající průchodu iontů H+ k syntéze ATP, nepatří proto vyloženě mezi primární aktivní transportéry.
- ABC transportéry
Tyto ATPázy představují obrovské množství různých transmembránových proteinů schopných aktivně přenášet různé látky dovnitř či ven přes membránu za spotřeby ATP. Mohou přenášet sacharidy, aminokyseliny, lipidy, ionty, peptidy i proteiny, toxiny a léky včetně antibiotik a další.[11]
Sekundární aktivní transport
Nazývá se také kotransport nebo spřažený přenašeč. Využívá energii přenosu určité látky ve směru jejího koncentračního gradientu k aktivnímu přenosu jiné látky proti směru jejího koncentračního gradientu. Koncentrační gradient první látky byl předtím vytvořen na jiném místě primárním aktivním transportem. Podle směru přenosu se rozlišuje symport a antiport.[12]
Symport
Při symportu jsou obě látky přenášeny přes membránu stejným směrem.
Příkladem je přenašeč glukosy a sodíku (SGLT1) v tenkém střevě, kdy glukosa je transportována z lumen střeva do enterocytu proti koncentračnímu gradientu a zároveň je do enterocytu resorbován sodík po koncentračním gradientu. Jde tedy o důležitý článek v metabolismu glukosy. Gradient pro sodík je vytvářen Na+/K+-ATPázou v jiné části membrány.[13][14] Dalším příkladem je GABA transportér spřažený s přenosem Na+ a Cl−, který zajišťuje hladinu neurotransmiteru GABA v synaptických štěrbinách.[15] Také přenos pyruvátu společně s H+ do mitochondrií je symportem.[9]
Antiport
Při antiportu se přenášené látky pohybují každá v opačném směru.
Takto funguje například výměník Ca2+/Na+, který hlavně v buňkách srdeční svaloviny udržuje nízkou hladinu vápenatých iontů uvnitř. Ca2+ je tedy transportován ven výměnou za tři Na+.[16] Zajímavostí je, že tento proces lze za určitých podmínek převrátit. Dalším příkladem je sekrece vodíkových iontů v ledvinných tubulech, kdy vodík je transportován z ledvinné buňky do lumen tubulu proti koncentračnímu gradientu a zároveň je do buňky resorbován sodík po koncentračním gradientu. Gradient pro sodík je vytvářen Na+/K+-ATPázou v jiné části membrány.[17]
Vezikulární transport
Zvláštním případem aktivního transportu je endocytóza a exocytóza, kdy jsou větší polární molekuly nebo skupina látek transportovány dovnitř nebo ven z buňky pomocí vezikulů (membránových váčku).[7]
Odkazy
Reference
V tomto článku byl použit překlad textu z článku Active transport na anglické Wikipedii.
- Homeostasis: keeping the body in balance. [s.l.]: Routledge Dostupné online. ISBN 978-1-315-84760-3. S. 23–30.
- DU BOIS-REYMOND, Emil. Untersuchungen über thierische Elektricität. Annalen der Physik und Chemie. 1848, roč. 151, čís. 11, s. 463–464. Dostupné online [cit. 2021-01-20]. ISSN 0003-3804. DOI 10.1002/andp.18481511120.
- ROSENBERG, Thomas; JOHANSON, Monica. On Accumulation and Active Transport in Biological Systems. I. Thermodynamic Considerations.. Acta Chemica Scandinavica. 1948, roč. 2, s. 14–33. Dostupné online [cit. 2021-01-20]. ISSN 0904-213X. DOI 10.3891/acta.chem.scand.02-0014. (anglicky)
- SKOU, Jens C. Die Identifizierung der Natrium-Kalium-Pumpe (Nobel-Vortrag). Angewandte Chemie. 1998-09-04, roč. 110, čís. 17, s. 2452–2461. <2452::aid-ange2452>3.0.co;2-z Dostupné online [cit. 2021-01-20]. ISSN 0044-8249. DOI 10.1002/(sici)1521-3757(19980904)110:17<2452::aid-ange2452>3.0.co;2-z.
- KOTYK, A. Symposium on Membrane Transport and Metabolism. Folia Microbiologica. 1961-03, roč. 6, čís. 2, s. 141–143. Dostupné online [cit. 2021-01-20]. ISSN 0015-5632. DOI 10.1007/bf02868237.
- UZMAN, A. Molecular Cell Biology (4th edition) Harvey Lodish, Arnold Berk, S. Lawrence Zipursky, Paul Matsudaira, David Baltimore and James Darnell; Freeman & Co., New York, NY, 2000, 1084 pp., list price $102.25, ISBN 0-7167-3136-3. Biochemistry and Molecular Biology Education. 2001, roč. 29, čís. 3, s. 126–128. Dostupné online [cit. 2021-01-20]. ISSN 1470-8175. DOI 10.1016/s1470-8175(01)00023-6.
- REECE, JANE B. Campbell biology. Tenth edition. vyd. Boston: [s.n.] 1 volume (various pagings) s. Dostupné online. ISBN 978-0-321-77565-8, ISBN 0-321-77565-1. OCLC 849822337
- COOPER, GEOFFREY M. The cell : a molecular approach. 5. vyd. Washington, D.C.: ASM Press xix, 820 pages s. Dostupné online. ISBN 978-0-87893-300-6, ISBN 0-87893-300-X. OCLC 310075904
- KODÍČEK, Milan. Biochemické pojmy - výkladový slovník [online]. Vydavatelství VŠCHT Praha, 2007. Dostupné online.
- FEHER, Joseph. Quantitative Human Physiology An Introduction. Second Edition. vyd. [s.l.]: Academic Press, 2017. ISBN 978-0-12-800883-6. S. 170–181.
- GOFFEAU, André; DE HERTOGH, Benoît; BARET, Philippe V. ENCYCLOPEDIA OF BIOLOGICAL CHEMISTRY, FOUR-VOLUME SET. [s.l.]: Elsevier Dostupné online. ISBN 978-0-12-443710-4.
- WAGNER, Hugh. Essentials of human physiology. Classic edition. | Abingdon, Oxon; New York, NY: Routledge, 2021. | Series: Psychology Press & Routledge Classics: Routledge Dostupné online. ISBN 978-1-003-14406-9. S. 11–24.
- DYER, J; HOSIE, K B; SHIRAZI-BEECHEY, S P. Nutrient regulation of human intestinal sugar transporter (SGLT1) expression.. Gut. 1997-07-01, roč. 41, čís. 1, s. 56–59. Dostupné online [cit. 2021-01-21]. ISSN 0017-5749. DOI 10.1136/gut.41.1.56. PMID 9274472. (anglicky)
- WRIGHT, E. M.; HIRAYAMA, B. A.; LOO, D. F. Active sugar transport in health and disease. Journal of Internal Medicine. 2007, roč. 261, čís. 1, s. 32–43. Dostupné online [cit. 2021-01-21]. ISSN 1365-2796. DOI 10.1111/j.1365-2796.2006.01746.x. (anglicky)
- Glutamate and gaba receptors and transporters : structure, function and pharmacology. London: Taylor & Francis xii, 435 pages s. Dostupné online. ISBN 0-7484-0881-9, ISBN 978-0-7484-0881-8. OCLC 48237833
- YU, Shan Ping; CHOI, Dennis W. Na+—Ca2+ Exchange Currents in Cortical Neurons: Concomitant Forward and Reverse Operation and Effect of Glutamate. European Journal of Neuroscience. 1997, roč. 9, čís. 6, s. 1273–1281. Dostupné online [cit. 2021-01-21]. ISSN 1460-9568. DOI 10.1111/j.1460-9568.1997.tb01482.x. (anglicky)
- KLOUDA, PAVEL. Základy biochemie. 2., přeprac. vyd. vyd. Ostrava: Pavel Klouda 143 s. Dostupné online. ISBN 80-86369-11-0, ISBN 978-80-86369-11-2. OCLC 85162209