Replikace DNA
Replikace DNA je proces tvorby kopií molekuly deoxyribonukleové kyseliny (DNA), čímž se genetická informace přenáší z jedné molekuly DNA (templát, matrice) do jiné molekuly stejného typu (tzv. replika). Celý proces je semikonzervativní, tzn. každá nově vzniklá molekula DNA má jeden řetězec z původní molekuly a jeden nový, syntetizovaný. Při replikaci dochází pomocí složité enzymatické mašinérie k řazení deoxyribonukleotidů (nukleových bází, základních stavebních částic DNA) jeden za druhým, a to podle vzorové původní molekuly DNA. Výsledkem tohoto řazení nukleotidů je nakonec kompletní DNA daného organizmu, v podstatě identická kopie původní DNA.
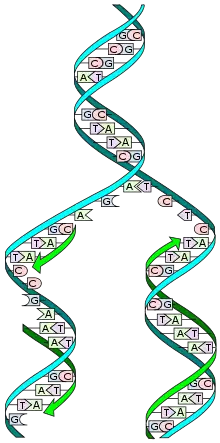
Do tajů tohoto procesu začali vědci blíže pronikat až v 60. a 70. letech minulého století a dodnes byly odhaleny do poměrně velkých podrobností molekulární pochody, jež se při replikaci odehrávají. Je například známo, že u bakterií probíhá replikace poněkud odlišně, než u eukaryotických organizmů, jako je například člověk, rostliny či houby. Praktický význam mají výzkumné metody odvozené od procesu replikace, jako je PCR a sekvenování.
Historie výzkumu
.jpg.webp)
Arthur Kornberg objevil v roce 1957 při studiu bakterie Escherichia coli první DNA polymerázu, jež nese jméno DNA polymeráza I.[1][2] Ačkoliv je dnes známo, že hlavní roli má v replikaci spíše DNA polymeráza III, i tak je tento objev důležitým milníkem. V roce 1958 bylo zjištěno, že replikace probíhá tzv. semikonzervativně (více viz Meselsonův-Stahlův experiment). V šedesátých letech byl učiněn další krok kupředu, když bylo zjištěno, že každé z vláken původní DNA je replikováno mírně odlišným způsobem, u jednoho z nich dochází vlivem diskontinuální syntézy ke vzniku Okazakiho fragmentů.[3]
Regulace a načasování
Načasování DNA replikace je diametrálně odlišné při srovnávání tohoto jevu u prokaryot a eukaryot. U bakterií a archeí (souhrnně prokaryota) nedochází k replikaci ve speciální fázi buněčného cyklu a nemusí vůbec souviset s buněčným dělením. Zato u eukaryot se obvykle replikace odehrává pouze v tzv. S fázi („S“ podle slova „syntéza“) buněčného cyklu. Mimo S fázi se syntéza DNA omezuje na drobné opravné mechanismy. Ve vzácných výjimkách (především tzv. endoreduplikace) však může i u eukaryot docházet k replikaci DNA bez ohledu na buněčný cyklus. Příkladem jsou slinné žlázy octomilky (Drosophila), v nichž se mnohonásobně nakopíruje genom, ačkoliv k replikaci nedojde.[4]
Parametry
Chybovost replikace DNA byla (u bakterií) odhadnuta na jednu chybu za 109–1010 nukleotidů (po proběhlém proofreadingu a korekci párování bází, tzv. mismatch repair).[5] Oprava DNA je zřejmě důvod, proč pracuje DNA polymeráza pouze v jednom směru (5'—>3'), v opačném směru by po opravě špatně začleněného nukleotidu nemohla pokračovat elongace řetězce DNA.[3]
Rychlost replikace genomu je u Escherichia coli asi 50 000 bází za minutu, zatímco u eukaryot je mnohem nižší. U pivní kvasinky činí 3 600 bází za minutu, u myši pouze 2 200 bází za minutu. Je tedy vidět, že ačkoliv mají eukaryota více replikačních počátků (kvasinka 400, myš kolem 2 500), je přesto tempo replikace jejich genomu pomalejší.[4] Kdyby byl u eukaryot k dispozici pouze jeden replikační počátek, trvalo by neúnosně dlouho, než by se celý genom replikoval: u člověka totiž rychlost DNA polymerázy činí pouhých 50 nukleotidů za sekundu.[6] Celý lidský genom (~3,2 mld. párů bází = ~3,2 Gb) by se tak z jediného replikačního počátku replikoval ~740 dní.
Obecný průběh
Při replikaci DNA vznikají v typickém případě z jedné dvoušroubovice DNA dvě tyto dvoušroubovice. Původní DNA se označuje jako matrice či templát, nově vytvořená DNA je někdy označována jako replika.
Replikace je přitom semikonzervativní, což znamená, že každý nový DNA řetězec se skládá z jednoho původního vlákna a jednoho nového vlákna. Do celého procesu je zapojeno mnoho enzymů, jako je především DNA polymeráza, dále však např. DNA ligáza, DNA primáza, helikáza či topoizomeráza. Každé z vláken původní dvoušroubovice je replikováno odlišným způsobem, což je způsobeno tím, že je molekula DNA tzv. antiparalelní. Zjednodušeně řečeno, skládá se ze dvou vláken, každé však je orientováno opačným směrem. DNA polymeráza však dokáže pracovat pouze v jednom směru (5'—>3'), což je v buňce vyřešeno elegantním způsobem. V každém případě však dochází k tomu, že je podle templátu (původní DNA) vytvářena nová DNA, která je k původnímu řetězci komplementární. To znamená, že když je v původní DNA detekována báze adenin, je do nové DNA přidán nukleotid obsahující thymin (a naopak, A a T k sobě patří). Pokud byl na templátové DNA nalezen například guanin, je do nového řetězce přidán podle pravidel komplementarity cytosinový nukleotid.[1]
Replikace je v základních rysech stejná u všech organizmů[3] a obecně je možné její průběh rozdělit do tří základních kroků:
- Iniciace – rozpletení dvoušroubovice DNA, vznik replikační vidlice a navázání enzymatického komplexu
- Elongace – přidávání nukleotidů a postup replikační vidlice
- Terminace – ukončení replikace
Iniciace
Replikace nezačíná na náhodném místě genomu, naopak toto místo je přesně určeno a označuje se jako replikační počátek. Některé organizmy těchto počátků mají několik tisíc (a tak vedle sebe při replikaci probíhá několik nezávislých polymerací), jednodušším organizmům s menším genomem stačí někdy pouze jeden replikační počátek.
K replikaci je na počátku potřeba krátká molekula RNA o délce 10–200 nukleotidů, tzv. primer.[1] Tuto sekvenci vytváří speciální enzym RNA-polymerizační označovaný jako primáza.[3] Primer se sice později odstraní a nahradí DNA, ale hraje významnou roli v iniciaci replikace. Celý replikační proces totiž začíná nukleofilní atakou 3'-hydroxylové skupiny tohoto primeru na fosfátovou skupinu prvního deoxyribonukleotidu. Dále se již deoxyribonukleotidy vážou na 3' uhlík předchozího deoxyribonukleotidu a nikoliv na primer; ten tedy slouží jen jednorázově[1] (to se však netýká opožďujícího se řetězce, viz kapitola „elongace“).
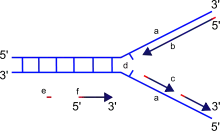
Vzniku replikační vidlice se účastní hexamerické enzymy helikázy, schopné oddálit obě molekuly dvoušroubovice a vytvořit charakteristickou strukturu ve tvaru písmene Y, aby mezi nimi vznikl prostor pro syntézu DNA. Udržet vlákna rozdělená pak pomáhají proteiny SSB proteiny. Helikázy však sice oddálí obě vlákna, ale dvoušroubovice se na konci replikační vidlice čím dál tím více utahuje a vzniká zde příliš velký tlak. Tento problém řeší topoizomerázy, jež jsou schopné přestřihnout jedno z vláken, uvolnit tlak a posléze ho opět slepit.[3][4]
Elongace
Poté, co je rozvinuta DNA a na vlákna přisednou DNA polymerázy, je možné začít samotnou replikaci. Aby v průběhu této činnosti DNA polymeráza pevně držela na svém místě, pomáhají jí k tomu tzv. svorkové proteiny (tvořící posuvnou svorku, DNA clamp).[3]
Důležité je upozornit, že každé z vláken původní templátové DNA je replikováno odlišným způsobem. Důvodem je skutečnost, že je DNA polymeráza schopná pracovat jen ve směru 5'—>3', tedy od pátého uhlíku deoxyribózy. Tento enzym totiž umí připojit nové nukleotidy pouze na 3' uhlík deoxyribózy. Toto omezení poněkud komplikuje celý proces, protože DNA je antiparalelní, tedy jeden řetězec směřuje 5'—>3' směrem, ale druhý 3'—>5' směrem. Buňky však tento problém řeší elegantně. Po jednom řetězci (tzv. vedoucí řetězec, čili angl. leading strand) postupuje DNA polymeráza zcela běžným způsobem. Druhý řetězec, který je orientován ve směru 3'—>5', se označuje jako opožďující se řetězec (angl. lagging strand). Tento řetězec však musí být kopírován rovněž v 5'—>3' směru (jinak to buňky neumí), a tak DNA polymeráza replikuje tento řetězec po malých částech, tzv. Okazakiho fragmentech o délce asi 100–200 nukleotidů (ale asi 10krát delší jsou tyto fragmenty u prokaryot). Na začátku každého Okazakiho fragmentu se musí pokaždé vytvořit nový RNA primer, ten je později vystřihnut a Okazakiho fragmenty jsou pospojovány DNA ligázou do souvislého vlákna, jež je od vedoucího (leading) řetězce k nerozeznání.[3] Kvůli zvláštní replikaci na zpožďujícím se vlákně se také replikace označuje jako semidiskontinuální.[7]
Terminace
Replikace končí tehdy, když je zhotovena kopie celé DNA. U bakterií se v tento okamžik sejdou obě replikační vidlice a splynou spolu, u eukaryot splývají replikační vidlice tehdy, když dosyntetizují „svou“ část genomu. Protože jsou však eukaryotické chromozomy lineární, DNA polymerázy nejsou schopné replikovat jejich koncové části, tzv. telomery, a tak je replikovaná DNA nepatrně kratší, než původní. Po určitém počtu buněčných dělení by to začalo vadit (Hayflickův limit); v praxi to však nepředstavuje problém, protože během meiózy se velikost telomer obnovuje pomocí enzymů telomeráz.
Rozdíly v průběhu replikace
U prokaryot
Replikace u prokaryot je relativně dobře prozkoumaná, protože bakterie (a jejich menší genom) představují snadnější model než komplexní eukaryotické organizmy. Zásadní pro průběh replikace u prokaryot je skutečnost, že obvykle mají kruhovou molekulu DNA, tzv. nukleoid.
Bakterie Escherichia coli má jediný replikační počátek, a to v genovém lokusu nazvaném oriC. Z tohoto místa se pohybují oběma směry replikační vidlice, v nichž dochází k syntéze nových řetězců DNA podle vzoru původních řetězců. Hlavní polymerázou bakterií je DNA polymeráza III obsahující velké množství různých podjednotek. Polymerázy I a II hrají pouze méně významnou roli.[4]
Protože z lokusu oriC byly vyslány dvě replikační vidlice po kruhové molekule DNA, tyto vidlice se logicky potkají přibližně „na půl cesty“ kolem dokola. Asi 100 kB od tohoto středu se nachází terminační (ukončovací) oblasti o délce 23 párů bází. Tyto terminační sekvence existují celkem čtyři (dvě na každém z vláken dvoušroubovice) a označují se TerA, TerB, TerC a TerD. Tyto sekvence rozeznává protein Tus, schopný ukončit replikaci v momentu, kdy se replikační vidlice dostane k terminačním sekvencím.[4]
Replikace mitochondriální DNA a plastidové DNA vykazuje značnou podobnost s replikací DNA u prokaryot.[4][8] Zato archebakterie (Archaea), jež jsou řazeny mezi prokaryota, vykazují poměrně značné rozdíly ve srovnání s replikací bakterií. Zatímco u bakterií replikace probíhá jen z jednoho replikačního počátku, u archeí je těchto míst zpravidla více (podobně jako u eukaryot). Také příslušné DNA polymerázy jsou podobné spíše eukaryotním DNA polymerázám.[9]
U eukaryot
Eukaryotické genomy jsou obecně větší než prokaryotické, čemuž musí být celý proces replikace přizpůsoben. Na rozdíl od bakterií, jež mají pouze jeden replikační počátek (místo, odkud začíná replikace), mají eukaryota replikačních počátků více. U kvasinky Saccharomyces cerevisiae se tyto replikační počátky označují ARS a je jich kolem 400 (u obojživelníků však jich je až 15 000). Z těchto replikačních počátků směřují oběma směry replikační vidlice ve tvaru písmene Y. Když se potkají dvě protijdoucí replikační vidlice, jednoduše dojde k jejich splynutí.[4]
V replikaci eukaryotického genomu zřejmě hraje roli větší množství různých proteinů. Příkladem je situace u SSB proteinů: u bakterií jsou tyto „stabilizátory jednovláknové DNA“ složené z jediné podjednotky, u eukaryot se skládají ze tří podjednotek.[3] Situace je také komplikovanější, co se týká počtu DNA polymeráz. U eukaryot jich bylo nalezeno nejméně 15.[10] DNA polymeráza α obsahuje podjednotku, jež funguje jako primáza, a je schopná vytvořit na začátku každého Okazakiho fragmentu RNA primer, načež k němu přidá několik DNA nukleotidů. Poté zřejmě předá místo na 3' konci prodlužujícího se řetězce DNA polymeráze δ. Vedoucí řetězec je zřejmě rovněž načat DNA polymerázou α, ale na rozdíl od opožďujícího se řetězce se zde zřejmě předává vlákno DNA polymeráze ε (ale stále o tom panují diskuse).[11][12]
U virů
Viry, které stojí na pomezí živého a neživého, mají poměrně specifický typ replikace DNA, odvíjející se od jejich parazitického způsobu života. Toto se netýká tzv. RNA virů, protože u většiny z nich v jejich rozmnožovacím cyklu molekuly DNA vůbec nefigurují (u těch ostatních, např. u retrovirů, molekula DNA figuruje, ale nedochází k její replikaci DNA polymerázou). Replikace DNA se tedy týká výhradně DNA virů. Některé DNA viry obsahují jednovláknovou DNA, a tak po infekci musí nejprve dojít k syntéze komplementárního vlákna, aby vznikla klasická dvoušroubovice. U virů se vyskytuje poměrně velké množství dalších odchylek od běžného schématu DNA replikace.[4] Co se týče Polyomaviru, jenž infikuje eukaryotické buňky, jeho DNA je replikována téměř stejně jako vlastní eukaryotický genom. Důvod je prostý: viry často zneužívají hostitelské DNA polymerázy a celou jejich enzymatickou mašinérii. Jediný rozdíl spočívá v tom, že u Polyomaviru musí dojít k navázání tzv. velkého T antigenu (large T-antigen) na replikační počátek.[13]
Využití ve výzkumu
Přirozený mechanismus replikace DNA využívají mnohé metody moderní molekulární biologie a genetiky. Je na tom založena například polymerázová řetězová reakce (PCR), schopná namnožit (amplifikovat) vzorek DNA. Také mnohé způsoby sekvenování zahrnují replikaci daného vzorku DNA; příkladem je Sangerova metoda či pyrosekvenování.
Odkazy
Reference
- MURRAY, Robert K., et al. Harperova biochemie, z angl. 23. vyd.. . 4. vyd. v ČR.. vyd. Praha: H & H,, 2002. ISBN 80-7319-013-3.
- Lehman, I. R., Bessman, M. J.; Simms, E. S.; Kornberg, A. Enzymatic Synthesis of Deoxyribonucleic Acid. I. Preparation of Substrates and Partial Purification of an Enzyme from Escherichia coli. J. Biol. Chem.. July 1958, roč. 233, čís. 1, s. 163–170. Dostupné online. PMID 13563462.
- Alberts, Bruce , et al. The Molecular Biology of the Cell. [s.l.]: Garland Science, 2002. (4th. ed). Dostupné online. ISBN 0-8153-3218-1. Kapitola DNA Replication Mechanisms.
- RÉDEI, George P. Encyclopedia of Genetics, Genomics, Proteomics, and Informatics. 3rd Edition. vyd. [s.l.]: Springer, 2008. ISBN 978-1-4020-6753-2.
- ALBERTS, Bruce, et al. Essential Cell Biology. 2. vyd. New York: Garland Science, 2004. Dostupné online.
- Speed of DNA Replication [online]. Dostupné online.
- Okazaki and Semidiscontinuous Replication [online]. Facultad de Biología; Universidad de La Habana [cit. 2011-05-04]. Dostupné v archivu pořízeném dne 2012-04-11.
- MORI, Yoko, Seisuke Kimura, Ai Saotome, Nobuyuki Kasai, Norihiro Sakaguchi, Yukinobu Uchiyama, Toyotaka Ishibashi, Taichi Yamamoto, Hiroyuki Chiku, Kengo Sakaguchi. Plastid DNA polymerases from higher plants, Arabidopsis thaliana. Biochemical and Biophysical Research Communications. 2005-08-19, roč. 334, čís. 1, s. 43–50. Dostupné online. ISSN 0006-291X. DOI 10.1016/j.bbrc.2005.06.052.
- Kelman L. M., Kelman Z. Multiple origins of replication in archaea. Trends Microbiol.. 2004, roč. 12, čís. 9, s. 399–401. DOI 10.1016/j.tim.2004.07.001. PMID 153371581.
- Tuusa. Human DNA polymerase ε; Expression, phosphorylation and protein-protein interactions: Eukaryotic DNA polymerases [online]. Dostupné online.
- Pursell, Z.F. et al. Yeast DNA Polymerase ε Participates in Leading-Strand DNA Replication. Science. 2007, roč. 317, s. 127–130. DOI 10.1126/science.1144067. PMID 17615360.
- Scott D McCulloch; Thomas A Kunkel. The fidelity of DNA synthesis by eukaryotic replicative and translesion synthesis polymerases. Cell Research. 01/2008, roč. 18, s. 148–161. DOI 10.1038/cr.2008.4. PMID 18166979.
- VIROLOGY - CHAPTER THREE; DNA VIRUS REPLICATION STRATEGIES [online]. University of California. Dostupné online.
Související články
Externí odkazy
 Obrázky, zvuky či videa k tématu replikace DNA na Wikimedia Commons
Obrázky, zvuky či videa k tématu replikace DNA na Wikimedia Commons - (anglicky) Flash animace – replikace DNA
- (anglicky) Další animace zachycující replikační vidlici
- (anglicky) Animované video s tematikou replikace DNA (k dispozici i na YouTube)
- (anglicky) Vtipnou formou podaná DNA replikace – text a obrázky
- (anglicky) Odpovídající kapitola v knize „Molecular Biology of the Cell“
- (anglicky) Video "DNA Replication Process" na YouTube