Zkřížená prezentace
Zkřížená prezentace (anglicky cross-presentation) je proces u antigen prezentujících buněk, při kterém dochází k vazbě exogenního antigenu nikoli na MHC II, ale na MHC I komplex a tím jeho prezentaci cytotoxickým CD8+ T lymfocytům (CD8+ Tc). Tento mechanismus se hlavně uplatňuje v antivirové imunitní odpovědi vůči virem infikovaným buňkám, které jsou tak díky zkřížené prezentaci cíleně identifikovány a zničeny.
Klasická vs. zkřížená prezentace
Principem MHC molekul je informovat imunitní systém o činnosti uvnitř buněk a složení extracelulárního prostředí. Každá jaderná buňka exprimuje na svém povrchu MHC I a prezentuje tak část svého vnitřního obsahu. Pohlcené antigeny z extracelulárního prostředí jsou prezentovány na MHC II, které exprimují hlavně antigen prezentující buňky. Tímto rozdělením intracelulární/extracelulární rozumíme klasickou antigenní prezentaci, která platí u většiny buněk.[1] Zkříženou prezentaci pozorujeme hlavně u dendritických buněk, které prezentují i exogenní antigeny na MHCI a tímto mechanismem tak aktivují CD8+ Tc cíleně namířené proti infikovaným buňkám.[2][3]
Mechanismus
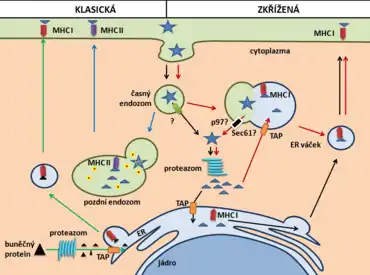
Po fagocytóze exogenního proteinu (například virového proteinu) jsou zde dvě možnosti, jak jeho fragmenty dostat na MHC I.
První a pravděpodobně nejčastější možností je tzv. cytosolická dráha. Zde je endocytovaný protein transportován z časného endozomu do cytoplazmy. Následuje štěpení v proteazomu na malé fragmenty a jejich transport do endoplazmatického retikula (ER) pomocí TAP transportéru. Fragmenty jsou poté navázány na MHC I a váčkem dopraveny a vystaveny na cytoplazmatické membráně. Obdobně může dojít ke splynutí časného endozomu s váčkem odvozeným od ER a transportu proteinu do cytoplazmy. Protein je tak opět naštěpen proteazomem a fragmenty jsou pak transportovány přes TAP zpět do endozomu (nebo do ER), kde se naváží na MHC I a jsou váčkem přeneseny a vystaveny na membráně.[4][5] Klíčovou roli tu tak mají TAP a proteazom, jejichž inhibice zruší nejen zkříženou prezentaci, ale i klasickou MHC I prezentaci.[6][7]
Přesný mechanismus transportu proteinu z endozomu do cytoplazmy doposud nebyl zcela jednoznačně objasněn. Jednou z možností je role Sec61 translokonu a p97/VCP, jakožto součást ERAD dráhy. Oboje se však vyskytují na membráně ER a od něj odvozených váčků. Vysvětloval by tak pouze transport proteinu po fúzi endozomu s ER váčkem.[8][9][10]
Druhou možností je tzv. vakuolární dráha. U té dochází k endocytóze proteinu a následné fúzi endozomu s lyzozomálnímí a ER váčky. Tato cesta je nezávislá na proteazomu a TAP transportéru, jelikož je protein degradován na fragmenty katepsinem S přímo ve váčku. Tyto fragmenty jsou poté přímo navázány na MHC I a vystaveny na cytoplazmatické membráně.[11][4]
Role v imunitní odpovědi
Zkřížené prezentace jsou schopné i makrofágy[12] a B lymfocyty[13], avšak v největší míře ji využívají právě dendritické buňky, konkrétně plazmacytoidní dendritické buňky.[14][2] Tyto plazmacytoidní dendritické buňky jsou klíčoví hráči v antivirové obraně, nejen díky vysoké produkci interferonu, ale i díky aktivaci CD8+ Tc[15]. Kdyby u nich platila pouze klasická prezentace, CD8+ Tc by nemohly být aktivovány exogenním antigenem. Ten by byl vázán pouze na MHC II a na ten se CD8+ Tc neváže. Plasmacytoidní dendritická buňka tak musí extracelulární antigen zpracovat tak, aby byl prezentován na MHC I. Je-li vir schopen nakazit dendritickou buňku, přirozeně se jeho části proteinů objeví na MHC I, jelikož jsou syntetizovány uvnitř buňky a tudíž jsou prezentovány stejně jako buňce vlastní proteiny. V tomto případě není potřeba zkřížené prezentace pro aktivaci CD8+ Tc . Avšak ne všechny viry dokážou infikovat i dendritické buňky, většina má specifický buněčný typ, ve kterém je virus schopen se zreplikovat. Proto dendritické buňky využívají zkřížené prezentace. Tím je zaručena eliminace infikovaných buněk, které na MHC I exprimují virové fragmenty, a i šíření viru a progrese nemoci. Role zkřížené prezentace a CD8+ Tc není však pevně svázána pouze s viry[16], ale i s bakteriální infekcí[17] a nádorovými onemocněními.[18][19]
Reference
- MANTEGAZZA, Adriana R.; MAGALHAES, Joao G.; AMIGORENA, Sebastian. Presentation of phagocytosed antigens by MHC class I and II. Traffic (Copenhagen, Denmark). 2017-02-01, roč. 14, čís. 2, s. 135–152. PMID: 23127154 PMCID: PMC3538944. Dostupné online [cit. 2017-02-01]. ISSN 1398-9219. DOI 10.1111/tra.12026. PMID 23127154.
- TEL, Jurjen; SCHREIBELT, Gerty; SITTIG, Simone P. Human plasmacytoid dendritic cells efficiently cross-present exogenous Ags to CD8+ T cells despite lower Ag uptake than myeloid dendritic cell subsets. Blood. 2013-01-17, roč. 121, čís. 3, s. 459–467. PMID: 23212525. Dostupné online [cit. 2017-02-01]. ISSN 1528-0020. DOI 10.1182/blood-2012-06-435644. PMID 23212525.
- DELAMARRE, Lélia; HOLCOMBE, Hilda; MELLMAN, Ira. Presentation of Exogenous Antigens on Major Histocompatibility Complex (MHC) Class I and MHC Class II Molecules Is Differentially Regulated during Dendritic Cell Maturation. Journal of Experimental Medicine. 2003-07-07, roč. 198, čís. 1, s. 111–122. PMID: 12835477. Dostupné online [cit. 2017-02-01]. ISSN 0022-1007. DOI 10.1084/jem.20021542. PMID 12835477. (anglicky)
- ROCK, Kenneth L.; SHEN, Lianjun. Cross-presentation: underlying mechanisms and role in immune surveillance. Immunological Reviews. 2005-10-01, roč. 207, s. 166–183. PMID: 16181335. Dostupné online [cit. 2017-02-01]. ISSN 0105-2896. DOI 10.1111/j.0105-2896.2005.00301.x. PMID 16181335.
- GUERMONPREZ, Pierre; SAVEANU, Loredana; KLEIJMEER, Monique. ER-phagosome fusion defines an MHC class I cross-presentation compartment in dendritic cells. Nature. 2003-09-25, roč. 425, čís. 6956, s. 397–402. PMID: 14508489. Dostupné online [cit. 2017-02-01]. ISSN 1476-4687. DOI 10.1038/nature01911. PMID 14508489.
- CRAIU, A.; GACZYNSKA, M.; AKOPIAN, T. Lactacystin and clasto-lactacystin beta-lactone modify multiple proteasome beta-subunits and inhibit intracellular protein degradation and major histocompatibility complex class I antigen presentation. The Journal of Biological Chemistry. 1997-05-16, roč. 272, čís. 20, s. 13437–13445. PMID: 9148969. Dostupné online [cit. 2017-02-01]. ISSN 0021-9258. PMID 9148969.
- DAY, P. M.; ESQUIVEL, F.; LUKSZO, J. Effect of TAP on the generation and intracellular trafficking of peptide-receptive major histocompatibility complex class I molecules. Immunity. 1995-02-01, roč. 2, čís. 2, s. 137–147. PMID: 7895170. Dostupné online [cit. 2017-02-01]. ISSN 1074-7613. PMID 7895170.
- ZEHNER, Matthias; MARSCHALL, Andrea L.; BOS, Erik. The translocon protein Sec61 mediates antigen transport from endosomes in the cytosol for cross-presentation to CD8(+) T cells. Immunity. 2015-05-19, roč. 42, čís. 5, s. 850–863. PMID: 25979419. Dostupné online [cit. 2017-02-01]. ISSN 1097-4180. DOI 10.1016/j.immuni.2015.04.008. PMID 25979419.
- ACKERMAN, Anne L.; GIODINI, Alessandra; CRESSWELL, Peter. A role for the endoplasmic reticulum protein retrotranslocation machinery during crosspresentation by dendritic cells. Immunity. 2006-10-01, roč. 25, čís. 4, s. 607–617. PMID: 17027300. Dostupné online [cit. 2017-02-01]. ISSN 1074-7613. DOI 10.1016/j.immuni.2006.08.017. PMID 17027300.
- MÉNAGER, Jérémie; EBSTEIN, Frédéric; OGER, Romain. Cross-presentation of synthetic long peptides by human dendritic cells: a process dependent on ERAD component p97/VCP but Not sec61 and/or Derlin-1. PloS One. 2014-01-01, roč. 9, čís. 2, s. e89897. PMID: 24587108 PMCID: PMC3937416. Dostupné online [cit. 2017-02-01]. ISSN 1932-6203. DOI 10.1371/journal.pone.0089897. PMID 24587108.
- SHEN, Lianjun; SIGAL, Luis J.; BOES, Marianne. Important role of cathepsin S in generating peptides for TAP-independent MHC class I crosspresentation in vivo. Immunity. 2004-08-01, roč. 21, čís. 2, s. 155–165. PMID: 15308097. Dostupné online [cit. 2017-02-01]. ISSN 1074-7613. DOI 10.1016/j.immuni.2004.07.004. PMID 15308097.
- NORBURY, C. C.; HEWLETT, L. J.; PRESCOTT, A. R. Class I MHC presentation of exogenous soluble antigen via macropinocytosis in bone marrow macrophages. Immunity. 1995-12-01, roč. 3, čís. 6, s. 783–791. PMID: 8777723. Dostupné online [cit. 2017-02-01]. ISSN 1074-7613. PMID 8777723.
- HON, Huiming; ORAN, Alp; BROCKER, Thomas. B lymphocytes participate in cross-presentation of antigen following gene gun vaccination. Journal of Immunology (Baltimore, Md.: 1950). 2005-05-01, roč. 174, čís. 9, s. 5233–5242. PMID: 15843519. Dostupné online [cit. 2017-02-01]. ISSN 0022-1767. PMID 15843519.
- HOEFFEL, Guillaume; RIPOCHE, Anne-Claire; MATHEOUD, Diana. Antigen crosspresentation by human plasmacytoid dendritic cells. Immunity. 2007-09-01, roč. 27, čís. 3, s. 481–492. PMID: 17869134. Dostupné online [cit. 2017-02-01]. ISSN 1074-7613. DOI 10.1016/j.immuni.2007.07.021. PMID 17869134.
- SWIECKI, Melissa; COLONNA, Marco. The multifaceted biology of plasmacytoid dendritic cells. Nature Reviews. Immunology. 2015-08-01, roč. 15, čís. 8, s. 471–485. PMID: 26160613 PMCID: PMC4808588. Dostupné online [cit. 2017-02-01]. ISSN 1474-1741. DOI 10.1038/nri3865. PMID 26160613.
- SIGAL, L. J.; CROTTY, S.; ANDINO, R. Cytotoxic T-cell immunity to virus-infected non-haematopoietic cells requires presentation of exogenous antigen. Nature. 1999-03-04, roč. 398, čís. 6722, s. 77–80. PMID: 10078533. Dostupné online [cit. 2017-02-01]. ISSN 0028-0836. DOI 10.1038/18038. PMID 10078533.
- SVENSSON, M.; PFEIFER, J.; STOCKINGER, B. Bacterial antigen delivery systems: phagocytic processing of bacterial antigens for MHC-I and MHC-II presentation to T cells. Behring Institute Mitteilungen. 1997-02-01, čís. 98, s. 197–211. PMID: 9382741. Dostupné online [cit. 2017-02-01]. ISSN 0301-0457. PMID 9382741.
- ASANO, Kenichi; NABEYAMA, Ami; MIYAKE, Yasunobu. CD169-positive macrophages dominate antitumor immunity by crosspresenting dead cell-associated antigens. Immunity. 2011-01-28, roč. 34, čís. 1, s. 85–95. PMID: 21194983. Dostupné online [cit. 2017-02-01]. ISSN 1097-4180. DOI 10.1016/j.immuni.2010.12.011. PMID 21194983.
- GOLDSZMID, Romina S.; IDOYAGA, Juliana; BRAVO, Alicia I. Dendritic cells charged with apoptotic tumor cells induce long-lived protective CD4+ and CD8+ T cell immunity against B16 melanoma. Journal of Immunology (Baltimore, Md.: 1950). 2003-12-01, roč. 171, čís. 11, s. 5940–5947. PMID: 14634105. Dostupné online [cit. 2017-02-01]. ISSN 0022-1767. PMID 14634105.