Rodozměna
Rodozměna (metageneze, střídání generací) je v nejobecnějším smyslu označení pro životní cyklus různých eukaryotických organismů, v němž se střídají generace rozmnožující se nepohlavně s generacemi, které se rozmnožují pohlavně.[1] V rámci zoologie se takto mohou označovat životní cykly živočichů, kteří kromě sexuality využívají i pravidelné klonální pomnožování partenogenezí (např. perloočky), pučením (např. mnozí žahavci, salpy) či jinými způsoby. Jak pohlavně, tak nepohlavně se množící jedinci jsou v takových případech diploidní (2n, se dvěma sadami chromozomů v jádrech).
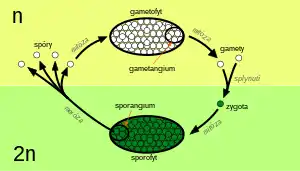
V rámci botaniky má pojem metageneze poněkud jiný, specifičtěji definovaný význam, neboť označuje takové životní cykly, kde nepohlavně se množící generace (sporofyt) je diploidní a meiózou produkuje spory, které jsou haploidní (n, s jedinou sadou chromozomů). Ze spor vyrůstá pohlavní generace (gametofyt) produkující mitózou gamety. Gamety pak splývají v diploidní zygotu, z níž roste nový sporofyt. Jde o tzv. haplo-diplontní (haplo-diploidní) cyklus, v rámci něhož nacházíme haploidní i diploidní mnohobuněčná stádia. To je principiální a biologicky relevantní rozdíl oproti živočichům, kde jediné haploidní buňky životního cyklu jsou zpravidla gamety (u některých skupin živočichů se vyskytují haploidní samci). Haplo-diplontní rodozměna je charakteristická pro všechny vyšší rostliny a mnohé řasy.
Alternativně je haplo-diplontní rodozměna definována jako životní cyklus, kde jak v haploidním, tak v diploidním stádiu probíhají mitotická dělení. Tuto mírně rozšířenou definici lze vztáhnout i na jednobuněčná eukaryota (prvoky), mezi kterými lze také haplo-diploidní cykly nalézt, např. u dírkonošců. I u nich se pak hovoří o rodozměně. Stejným pojmem se ale někdy označují i cykly prvoků, kde se jak pohlavní, tak nepohlavní množení uskutečňuje na stejné ploidní úrovni – např. v rámci „rodozměny” výtrusovců se sice střídá nepohlavní množení s tvorbou gamet, ale oboje se odehrává během haploidní části cyklu. Některými autory je za pravou metagenezi považována pouze rodozměna doprovázející haplo-diplontní životní cyklus.[2]
Zejména u řas se rozlišuje izomorfní rodozměna, kdy se gametofyt a sporofyt vzájemně podobají, a anizomorfní čili heteromorfní rodozměna, kdy se obě stádia zřetelně liší (to je i případ vyšších rostlin).
Rodozměna u rostlin
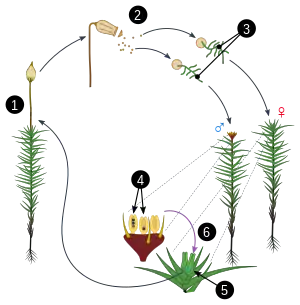
Vyšší rostliny
S klasickou metagenezí, kde se střídá haploidní a diploidní generace, tedy s tzv. haplo-diplontním cyklem,[3][2] se setkáváme u vyšších rostlin a u mnoha skupin řas.[4][5] V obecném případě vypadá metageneze rostlin následovně: mnohobuněčné diploidní stádium (sporofyt) ve výtrusnicích (sporangiích) různého charakteru pomocí meiózy produkuje haploidní jednobuněčné výtrusy (spory), které po úspěšném vyklíčení dále rostou a mitotickým dělením vytvářejí mnohobuněčné haploidní stádium (gametofyt). Gametofyt pak mitózou produkuje gamety v pohlavních orgánech (gametangiích), které lze rozlišit na samčí pelatky (anteridia) a samičí zárodečníky (archegonia). Gamety jsou podobně jako výtrusy jednobuněčné a haploidní, ale jejich rolí je splynutí, při němž vzniká diploidní zygota, ze které vyrůstá další sporofyt.[4] Metageneze tohoto typu je dosti zřetelná u výtrusných suchozemských rostlin (mechorostů a kapraďorostů), u nichž bývá sporofyt i gametofyt makroskopický. U semenných rostlin je již metageneze zastřenější a komplikovanější. Gametofyty jsou zde jednak odděleného pohlaví, jednak jsou silně redukované co do počtu buněk i co do velikosti (jsou mikroskopické). Samičí gametofyt semenných rostlin (zárodečný vak) je navíc trvale obklopen pletivy mateřského sporofytu.[4][6]
V rámci evoluce vyšších rostlin tak lze hovořit o redukci gametofytu a dominanci sporofytu: u mechorostů je gametofyt dominantním, samostatně žijícím fotosyntetickým stádiem, krátkověký sporofyt je na něm plně závislý. U výtrusných cévnatých rostlin (kapraďorostů) zůstává gametofyt samostatným organismem, ale je již výrazně drobnější než sporofyt, který přejímá roli dominantního stádia. Gametofyty semenných rostlin jsou již drasticky redukované, v případě krytosemenných až na pouhé 3 buňky (samčí gametofyt – klíčící pylové zrno) resp. na osmijaderný, sedmibuněčný útvar (samičí gametofyt – zárodečný vak).[4][7] U řas, které jsou fylogeneticky blízké vyšším rostlinám, jako jsou parožnatky nebo spájivky, sporofyt chybí – diploidní fáze je u nich zastoupená pouze zygotou, jejíž první dělení je meiotické. Jsou tak ukázkou haplontního cyklu. Mnohobuněčný sporofyt – a zároveň „pravá“ metageneze se střídáním obou mnohobuněčných fází – je pak evoluční novinkou (synapomorfií) vyšších rostlin (k níž ovšem konvergentně dospěli i zástupci dalších skupin).[8]
Řasy
U řas se (už kvůli jejich různorodosti dané jejich polyfyletickým původem) vyskytují nejrůznější typy životních cyklů. Kromě výše zmíněných haplontních organismů mezi nimi najdeme i takové, kde je jediným mnohobuněčným stádiem sporofyt a haploidní stádia jsou zastoupena pouze gametami. Takové organismy jsou označovány jako diplontní. Příkladem mohou být některé chaluhy, např. chaluha bublinatá.[9] Mnoho řas ale střídá gametofyt se sporofytem a jde tedy o haplo-diplonty. Podle některých definic je pouze v tomto případě možné mluvit o skutečném střídání generací.[2] Pokud se gametofyt a sporofyt haplo-diplontních řas vzájemně podobají a je obtížné je morfologicky od sebe odlišit, hovoří se o izomorfní rodozměně. Příkladem jsou např. porost locikový nebo žabí vlas. Gametofyt a sporofyt se ale od sebe mohou výrazně lišit, pak jde o anizomorfní čili heteromorfní rodozměnu (např. chaluha čepelatka prstnatá).[5] Heteromorfní rodozměna je typická i pro vyšší rostliny.[4]
Rodozměna u dalších eukaryot
Houby
U dikaryotických hub (vřeckatých a stopkovýtrusných) lze podobně jako u rostlin nalézt mnohobuněčná haploidní stádia, a pak mnohobuněčná stádia, jejichž buňky obsahují dvě sady chromozomů. Rozdíl spočívá v tom, že tyto dvě sady zůstávají ve dvou nesplývajících haploidních jádrech, proto se hovoří o dikaryotických buňkách, nejde o typické diploidní buňky.[6] U některých chytridiomycét (např. Allomyces) se skutečně střídá mnohobuněčné haploidní a diploidní stádium. Někdy se o nich hovoří jako o gametofytu a sporofytu, ale protože houby nejsou příbuzné rostlinám, vhodnější je nazývat je gametothallus a sporothallus.[10]
Prvoci
Termín „rodozměna“ je poněkud nekonzistentně používán u některých dalších skupin eukaryot. Někdy je užíván ve významu blízkém výše uvedeným variantám, označuje tedy situaci, kdy se rozmnožování odehrává jak na úrovni haploidní, tak i diploidní generace, které se v životním cyklu (více či méně pravidelně) střídají a často se od sebe i (více či méně nápadně) liší. Jde tedy o haplo-diplontní cykly definované v tomto případě tak, že jak v haploidní, tak v diploidní generaci probíhají mitotická dělení.[2][9] Tohoto charakteru je např. rodozměna plazmodiálních hlenek[11] nebo rodozměna dírkonošců.[12] Jindy se pojmem rodozměna označuje případ, kdy se v životním cyklu sice střídá fáze pohlavního a nepohlavního množení, obě se ale odehrávají na stejné ploidní úrovni. Tak např. „rodozměna“ výtrusovců zahrnuje intenzivní nepohlavní množení (merogonii neboli schizogonii) i tvorbu gamet (gamogonii), oboje se ale odehrává v haploidní fázi životního cyklu. Jediným diploidním stádiem je zde zygota, která se dělí meioticky (během tzv. sporogonie).[13] Jde tedy o haplontní cyklus odlišný od „typické“ haplo-diplontní rodozměny.
Živočichové
Životní cykly živočichů jsou prakticky vždy diplontní, jedinými haploidními buňkami jejich cyklu jsou gamety.[14] Jen výjimečně mohou u živočichů vznikat haploidní mnohobuněčná těla produkující gamety mitózou, a to haploidní samci u skupin s haplo-diploidním určením pohlaví (mnozí vířníci,[15] blanokřídlí,[16] aj.). Pojem „rodozměna“ se u živočichů používá tehdy (spíše však ve starší literatuře), pokud dochází během životního cyklu k výraznému střídání množení pohlavního a nepohlavního, ať už jde o partenogenezi (např. perloočky),[17] pučení (např. koloniální pláštěnci)[18] nebo jiné způsoby (strobilace medúz aj.).[19] Jedinci „nepohlavní“ a „pohlavní“ generace se však v tomto případě od sebe geneticky neliší, což je významný rozdíl proti situaci u „pravé“ rodozměny u rostlin.[20] Pro střídání partenogeneze se sexualitou se užívá pojem heterogonie.[21]
- Varianty rodozměny u eukaryot
.png.webp) Schéma rodozměny chytridiomycéty rodu Allomyces, kde se střídá haploidní gametothallus a diploidní sporothallus; M! symbolizuje ve schématu meiózu, P! splývání gamet (plazmogamii) a K! splývání jejich jader (karyogamii)
Schéma rodozměny chytridiomycéty rodu Allomyces, kde se střídá haploidní gametothallus a diploidní sporothallus; M! symbolizuje ve schématu meiózu, P! splývání gamet (plazmogamii) a K! splývání jejich jader (karyogamii).png.webp) Schéma rodozměny plazmodiálních hlenek, kde se mitózou dělí jak haploidní jádra (během asexuální fáze), tak i diploidní jádra (během růstu mnohojaderného plazmodia); vysvětlivky viz předchozí schéma
Schéma rodozměny plazmodiálních hlenek, kde se mitózou dělí jak haploidní jádra (během asexuální fáze), tak i diploidní jádra (během růstu mnohojaderného plazmodia); vysvětlivky viz předchozí schéma
Reference
- alternation of generations | Definition & Examples | Britannica. www.britannica.com [online]. [cit. 2021-12-27]. Dostupné online. (anglicky)
- INGROUILLE, Martin. Plants: evolution and diversity. Cambridge, UK: Cambridge University Press, 2006. 1 online resource (xiv, 440 pages) s. Dostupné online. ISBN 978-0-511-64852-6, ISBN 0-511-64852-9. OCLC 667094262 S. 31–34.
- KALINA, Tomáš. Systém a vývoj sinic a řas. Praha: Karolinum, 1998. S. 21.
- NOVÁK, František. Vyšší rostliny. 2. vyd. Praha: Academia, 1972. Kapitola Antithetická rodozměna, s. 41–49.
- KALINA, Tomáš. Systém a vývoj sinic a řas. Praha: Karolinum, 1998.
- KUBÁT, Karel, a kol. Botanika. [s.l.]: Scientia, 2003. ISBN 80-7183-266-9.
- SIMPSON, Michael G. Plant systematics. 1. vyd. Amsterdam: Elsevier/Academic Press, 2005. 1 online resource (xi, 590 pages) s. Dostupné online. ISBN 978-0-08-051404-8, ISBN 0-08-051404-9. OCLC 230198576
- SIMPSON, Michael G. Plant systematics. 1. vyd. Amsterdam: Elsevier/Academic Press, 2005. 1 online resource (xi, 590 pages) s. Dostupné online. ISBN 978-0-08-051404-8, ISBN 0-08-051404-9. OCLC 230198576 S. 53–54.
- HEESCH, Svenja; SERRANO‐SERRANO, Martha; BARRERA‐REDONDO, Josué. Evolution of life cycles and reproductive traits: Insights from the brown algae. Journal of Evolutionary Biology. 2021-07, roč. 34, čís. 7, s. 992–1009. Dostupné online [cit. 2021-12-26]. ISSN 1010-061X. DOI 10.1111/jeb.13880. (anglicky)
- WEBSTER, John. Introduction to fungi. 3rd ed. vyd. Cambridge, UK: Cambridge University Press, 2007. 1 online resource (xviii, 841 pages, 12 unnumbered pages of plates) s. Dostupné online. ISBN 978-0-511-64895-3, ISBN 0-511-64895-2. OCLC 593239922 Kapitola 6.5 Blastocladiales, s. 153–162.
- VÁŇA, Jiří. Systém a vývoj hub a houbových organismů. Praha: Karolinum, 1998. S. 19–21.
- HAUSMANN, Klaus; NORBERT, Hülsmann. Protozoologie. 1. vyd. Praha: Academia, 2003. ISBN 80-200-0978-7. S. 149.
- HAUSMANN, Klaus; NORBERT, Hülsmann. Protozoologie. 1. vyd. Praha: Academia, 2003. ISBN 80-200-0978-7. S. 100–102.
- HICKMAN, Cleveland P., Jr. Integrated principles of zoology. Eighteenth edition. vyd. New York, NY: [s.n.], 2020. 1 online resource s. Dostupné online. ISBN 1-260-56945-4, ISBN 978-1-260-56945-2. OCLC 1156618407 S. 140.
- Hickman, 2020, s. 317
- Hickman, 2020, s. 139
- HOLEC, Michael; HOLCOVÁ, Diana. Zoologie I. [s.l.]: Univerzita J. E. Purkyně v Ústí n. Labem, 2014. S. 43.
- ŠMIDT, G. A. Embryologie živočichů II: speciální embryologie. Praha: Nakladatelství ČSAV, 1960. S. 278.
- Šmidt , 1960, s. 40
- Bílý, M.; Hájek, J.; Koutecký, P.; Kratzerová, L. Rozmnožování organismů. Praha: Ústřední komise biologické olympiády, 2000. S. 19.
- BUCHAR, Jan, a kol. Klíč k určování bezobratlých. 1. vyd. Praha: Scientia, 1995. ISBN 80-85827-81-6. S. 252.