Šatovník šarlatový
Šatovník šarlatový (Drepanis coccinea) je druh pěnkavovitého ptáka ze skupiny šatovníků a rodu Drepanis, endemit Havajských ostrovů, jenž je rozšířen ve vlhkých a středně vlhkých lesích napříč hlavním řetězcem souostroví. Druh popsal Georg Forster roku 1780. Nejpočetnější populace stále přežívají na ostrově Havaji a na východním Maui (celkem více než 600 000 jedinců). Na ostrově Kauai je šatovník šarlatový omezen na náhorní plošinu Alakai, počty ptáků na Oahu a Molokai jsou nepatrné. Na Lanai šatovník šarlatový vyhynul již během první poloviny 20. století.
 Šatovník šarlatový | |
| Stupeň ohrožení podle IUCN | |
 zranitelný[1] | |
| Vědecká klasifikace | |
| Říše | živočichové (Animalia) |
| Kmen | strunatci (Chordata) |
| Třída | ptáci (Aves) |
| Řád | pěvci (Passeriformes) |
| Čeleď | pěnkavovití (Fringillidae) |
| Rod | šatovník (Drepanis) |
| Binomické jméno | |
| Drepanis coccinea (Forster, 1780) | |
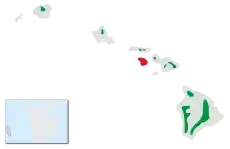 Rozšíření šatovníka šarlatového na Havajských ostrovech (červeně vyhynulé populace) Rozšíření šatovníka šarlatového na Havajských ostrovech (červeně vyhynulé populace)
| |
| Některá data mohou pocházet z datové položky. | |
Šatovník šarlatový dosahuje velikosti asi 15 cm. Vyznačuje se jásavě červeným zbarvením peří, s tmavými křídly a ocasem. Nedospělí jedinci mají peří především ve žlutých odstínech a konečné zbarvení se jim vyvine až v průběhu času. Pestré peří šatovníků bylo ceněno původním polynéským obyvatelstvem, které jej používalo jako součást honosných obleků vysoce postavených představitelů ostrovů. Výrazným znakem šatovníka šarlatového je také jeho prodloužený, dolů zahnutý zobák. Ten nejspíše představuje adaptaci ke konzumaci nektaru místních lobelek, s úbytkem těchto rostlin a přeorientováním se na jiné zdroje potravy se nejspíše postupně zkracuje a narovnává. Vyjma nektaru se šatovníci živí i bezobratlými živočichy, potravu sbírají především ve středních a vyšších patrech stromového baldachýnu. Vrchol rozmnožování je mezi únorem a červnem, do miskovitého hnízda samice snáší průměrně dvě vejce.
Mezinárodní svaz ochrany přírody považuje šatovníka šarlatového za zranitelný taxon. Hrozbu představuje především šíření ptačí malárie, parazitického onemocnění přenášeného nepůvodními druhy komárů, přičemž šatovníci šarlatoví jsou k infekcím obzvláště vnímaví a provází je vysoká úmrtnost. Vzhledem k tomu, že komáři žijí především v teplejších nížinných stanovištích, byli šatovníci šarlatoví v průběhu 20. století vytlačeni do výše položených areálů v nadmořských výškách nad 1 300 metrů. Budoucí hrozbu a výzvu pro ochránce přírody však představuje klimatická změna a s ní spojené rozšíření nemocí i do výše položených stanovišť.
Systematika
Objev a pojmenování

Šatovník šarlatový byl poprvé popsán v roce 1780 (ačkoli popis byl zveřejněn až v následujícím roce[2]), přičemž autorem popisu druhu byl Georg Forster, polsko-německý přírodovědec a cestovatel. Exempláře tohoto druhu Forster získal od Bartholda Lohmanna, jenž byl členem družiny Jamese Cooka při jeho poslední objevné plavbě, při níž byly pro západní civilizaci objeveny samotné Havajské ostrovy.[3][4] Sám Cook se zmiňuje, že místní sběratelé jim často dodávali svazky až dvaceti nebo více jedinců, nicméně nikdo z posádky tento druh neviděl živý (zřejmě proto, že se vyhýbal pobřežním oblastem).[2]
Forster zvolil pro šatovníka původní vědecké pojmenování Certhia coccinea, přičemž druhové jméno „coccinea“ (vycházející ze řeckého „κόκκινος“) odkazuje na šarlatové zbarvení peří. Následně byl šatovník šarlatový dlouhodobě řazen v rámci rodu Vestiaria, a to ještě během konce 20. a začátku 21. století. Moderní systematika jej společně se šatovníkem žlutoocasým (Drepanis pacifica) a šatovníkem černým (Drepanis funerea) sdružuje spíše do rodu Drepanis v rámci čeledi pěnkavovití a skupiny šatovníků (není zcela ustálena jejich taxonomická kategorie). V takovém případě představuje šatovník šarlatový jediný přežívající druh rodu, protože zbylé dva kongenerické druhy z volné přírody vymizely v důsledku lidských činností na přelomu 19. a 20. století.[5][3][4]
V havajštině nese šatovník šarlatový pojmenování „iiwi“, přičemž toto jméno je možná onomatopoion.[3] Jiným etymologickým vysvětlením je, že toto jméno vychází z pojmenování pro kolihu v malajsko-polynéských jazycích; zobák šatovníka šarlatového částečně připomíná zobák kolihy, takže připodobnění obou druhů je nasnadě.[6] Mimo obecného havajského jména existuje několik dalších lokálních pojmenování, jako „iawi“, „iiwipolena“, „ahikipolena“ (či zkráceně pouze „polena“). Na ostrově Kauai je nazýván „olokele“. Juvenilní zelenavá forma je nazývána „iiwipopolo“.[7]
Evoluce a příbuzenské vztahy

Šatovník šarlatový je původní na všech šesti největších ostrovech Havajského souostroví (na Lanai však vyhynul). Navzdory tomu se nezdá, že by se zde rozrůznil do specifických poddruhů; jako poddruh byl sice někdy hodnocen D. c. suavis z ostrova Molokai, nicméně tato systematika není široce uznávána. Je možné, že je tato situace výsledkem relativně nedávného osídlení jednotlivých ostrovů, stejně jako migrací jednotlivých šatovníků mezi ostrovy, díky čemuž nedochází k vytvoření účinných reprodukčních bariér.[3][4]
Nejbližšími žijícími příbuznými šatovníka šarlatového jsou šatovník chocholatý (Palmeria dolei) a šatovník karmínový (Himatione sanguinea).[4] Šatovník karmínový a šatovník šarlatový jsou schopni plodit i hybridní potomstvo, jak prokázala studie z roku 2014, jejíž autoři objevili hybridního jedince, který vznikl křížením samice šatovníka šarlatového a samce šatovníka karmínového. Jednalo se o první zaznamenaný hybrid v rámci celé početné skupiny šatovníků, třebaže hybridizace u nehavajských pěnkavovitých nejsou natolik výjimečné. Důvodem, proč se jinak evolučně nepříliš vzdálené druhy – stáří divergence mezi rodičovskými druhy bylo stanoveno na přibližně 1,6 milionu let – nekříží častěji, mohou být účinné reprodukčně-izolační mechanismy vzniknuvší významnými behaviorálními a morfologickými rozdíly. Zaznamenaný hybrid byl pozorován ve specifickém životním prostředí: šlo o lesní fragment, v němž byl šatovník karmínový několikanásobně hojnější než šatovník šarlatový, tj. k jevu mohla přispět vzácnost ptáka (omezená nabídka partnerů stejného druhu). Hypoteticky se může úroveň hybridizace mezi těmito dvěma druhy v budoucnu zvyšovat, protože šatovník šarlatový je ve srovnání se šatovníkem karmínovým extrémně náchylný k infekci ptačí malárií; kříženci šatovníka šarlatového tedy mohou být vůči tomuto onemocnění méně vnímaví.[8]
Původ samotných šatovníkovitých je pravděpodobně asijský, přičemž sesterským taxonem jsou hýlové z rodu Carpodacus. Nejranější divergence v rámci skupiny se kryjí se vznikem ostrovů Niihau (5,7–5,3 milionů let) a Kauai (5,4–4,9 milionů let), náležících k hlavnímu řetězci havajských ostrovů.[9] Šatovníkovití na Havaji pocházejí pravděpodobně z jediného předka. Následně prošli procesem masivní adaptivní radiace, rozšířili se skrze celé souostroví a začali se vyznačovat značnou mezidruhovou variabilitou; značné rozmanitosti dosahuje především tvar zobáků, které se pohybují od silných zobáků sloužících k pojídání semen až po protažené zobáky nektarožravých druhů, k nimž spadá i šatovník šarlatový.[10][11]
Popis

Šatovník šarlatový je středně velký druh šatovníka, dosahuje délky asi 15 cm a hmotnosti 16 až 20 g. Samec bývá o něco větší než samice, leč jinak není pohlavní dimorfismus příliš výrazný a obě pohlaví si jsou svým opeřením podobná.[11][12] Peří je na těle šarlatově-rumělkové, na bázi jsou pera šedavá. Křídla a ocas jsou zbarveny vesměs leskle černým opeřením. Barva krycích letek křídel se ale obvykle pohybuje i v určitých odstínech bílé a šedé, přičemž nejvnitřnější krycí letky bývají zcela bílé. Velké předloketní krovky křídel jsou úzce lemovány a zakončeny šarlatovou, dvě nejvnitřnější předloketní krovky jsou pak šarlatové s bílým lemováním a černým zakončením. Spodní strana křídel je matně bělavá, opět se šarlatovým nádechem; spodek ocasu je spíše žlutohnědě-šedavý. Celkové opeření, zvláště na světle, působí lesklým dojmem. Peří na hlavě se může lidskému pozorovateli zdát o něco světlejší než na zbytku těla, přičemž tento jev je zapříčiněn průběhem přepeřování šatovníka během roku. Peří na hlavě šatovník mění až na konci celého cyklu, kvůli čemuž vzniká kontrast mezi novými pery pokrývajícími tělo a starými pery na hlavě. Okolo oka je vytvořen kontrastující bledě oranžový kroužek.[13]

Šatovník šarlatový se vyznačuje dlouhým zahnutým zobákem, jenž dosahuje velikosti 25 až 28 mm a jeho zbarvení se pohybuje v broskvových a lososových odstínech.[12] Zobák slouží k pronikání do květů rostlin a vybírání nektaru. Existují náznaky, že s úbytkem místních druhů lobelek, jejichž nektarem se šatovníci živí, jejich specializovaný zobák prochází evolučními změnami a postupně se zkracuje a narovnává. Dochází tak k přizpůsobení se jiným zdrojům potravy, jako jsou květy druhu železnec mnohotvarý (Metrosideros polymorpha, havajsky ohia).[10]
Zbarvení nedospělých jedinců je spíše matně žluté a doplňují jej černé skvrnky. Juvenilní ptáci se liší také barvou zobáků, jež je tmavší než v případě dospělců.[12] V rámci jiných druhů šatovníků lze šatovníka šarlatového zaměnit s blízce příbuzným šatovníkem karmínovým. Šatovník karmínový se nicméně odlišuje bělavým zbarvením břicha a spodní strany ocasu, zatímco svrchní partie jsou vybarveny sytějšími odstíny červené. Významným rozlišovacím znakem je taktéž zobák, jenž je u šatovníka karmínového kratší a méně zahnutý, navíc má černé zbarvení.[11] Peří v červených odstínech některých havajských šatovníků může mít evoluční základ v podobnosti s barvou květů stromu ohia. Protože jde o důležitý potravní zdroj Havajských ostrovů, jenž poskytuje ptákům nektar i hmyz, rozvoj červeného opeření může vycházet z určitých samičích psychických predispozic, na základě kterých preferovaly samce o podobném zbarvení, jaké má tento klíčový druh dřeviny. Naopak se nezdá, že by červené opeření mělo sloužit jako maskování, protože na Havaji nežijí původní druhy predátorů, které by šatovníky nějak významně ohrožovaly.[14]
Šatovník šarlatový je dobrým letcem. Pera na křídlech vytvářejí během letu typický bzučivý zvuk, jenž je možné slyšet do vzdálenosti 50 až 100 metrů od letícího ptáka.[15]
Biologie
Výskyt a stanoviště
.jpg.webp)
Šatovník šarlatový je rozšířen skrze hlavní řetězec Havajských ostrovů, počty tohoto druhu však během 20. století značně poklesly. Největší populace stále přežívají na ostrovech Havaj, Maui a Kauai. Na Havaji šatovník šarlatový žije v pásmu táhnoucím se kolem velké části ostrova, izolovaná populace pak existuje na sopce Kohala na severu ostrova. Na Maui je šatovník šarlatový rozšířen primárně ve východních částech ostrova, malá populace ale obývá i západní Maui. Na Kauai je šatovník šarlatový omezen na náhorní plošinu Alakai. Malé reliktní izolované populace stále přežívají i na ostrovech Oahu a Molokai.[5] Historicky byl šatovník šarlatový hojný i na ostrově Lanai, nicméně do konce 20. let 20. století zde vyhynul.[7] Jde o zdatného letce, během roku mění stanoviště podle dostupnosti potravy.[4]
Šatovník šarlatový je druhem vlhkých a středně vlhkých lesů, občasně shání potravu i v suchých lesních stanovištích, nicméně nikdy zde nehnízdí. Havajským lesům dominují stromy ohia a koa (místní druh akácie), přičemž preferovaná vegetace bývá často doplněna i o další druhy místních rostlin, jako jsou kolea lau nui (druh lacháru), naio (vousatec havajský, Myoporum sandwicense) a stromové kapradiny ciboty. Ve výše položených stanovištích převládají porosty jerlínu zlatolistého (Sophora chrysophylla, havajsky mamane).[4] Historicky se šatovníci vyskytovali ve výškovém rozsahu od 300 do 2 900 metrů. Následkem dovlečení ptačí malárie přenášené komáry však byla drtivá většina populací vytlačena do výše položených stanovišť (1 300 metrů až 1 900 metrů) s chladnějším podnebím.[16]

Chování
Šatovník šarlatový se ozývá pestrou škálou vrzavých, pískavých a bublavých zvuků, některé tóny jsou připodobňovány hře na starou harmoniku, jiné rezavému pantu. Zpěv a volání jsou si velmi podobné. Šatovník může napodobovat zvuky jiného havajského ptactva, zejména místních lejskovců (Chasiempis sp.).[13] Obě pohlaví zpívají celoročně, ačkoli na začátku hnízdní sezóny se ptáci ozývají četněji. Zpěv šatovníků lze nejčastěji zaslechnout v brzkých ranních hodinách a během pozdního odpoledne.[15] Krmná teritoria si tento druh může bránit před ostatními druhy ptáků i proti jedincům vlastního druhu, přičemž agresivní chování snižuje husté olistění stromových korun. V rámci havajského ptactva je v současnosti na drtivé většině území dominantním druhem, jenž z krmných stanovišť vytlačuje ostatní původní opeřence. Velký vliv na tom má hromadné vymírání havajského ptactva a s ním spojený úbytek až úplné vymizení původních dominantních druhů.[17] Například britský přírodovědec Robert Cyril Layton Perkins, jenž žil mezi lety 1866 až 1955 a strávil velké množství času studiem divočiny souostroví, na začátku 20. století popisoval, jak byl pták moho ozdobný (Moho nobilis) z hlavního ostrova k šatovníkům šarlatovým velmi nesnášenlivý. Moho, jenž z ostrova zmizel do konce první poloviny 20. století, někdy dokonce „úplně opustil krmný strom, ať už byly jeho květy jakkoli bohaté, pouze aby vypudil z nějakého vzdáleného stromu šatovníka, jehož tam náhodou zahlédl“.[18] Jediným původním druhem, který je šatovníkovi šarlatovému nadřazen, je šatovník chocholatý (Palmeria dolei) na ostrově Maui, ač i tento pták je kriticky ohrožen vyhubením.[17]
.jpg.webp)
Šatovník šarlatový se živí především nektarem, v čemž mu napomáhá trubičkovitý jazyk s kartáčovitou špičkou, a jídelníček doplňuje i bezobratlými živočichy, jako jsou hmyz a pavouci.[11] Krmí se nejčastěji ve středních a vyšších patrech stromového baldachýnu, ačkoli někdy se může krmit i na nižších rostlinách. Naopak na zemi nesbírá potravu prakticky nikdy, na kmenech stromů jenom občasně. Na jednom stromě potravu hledá obyčejně samostatně, anebo v páru (a případně i s mláďaty).[19] Krmí se na místních lobelkách (rody Clermontia a Cyanea), stromech ohia (železnec), mamane (jerlín zlatolistý) i na nižších rostlinách, jako jsou místní ostružiníky (Rubus hawaiensis, havajsky akala) nebo keř kanawao (Broussaisia arguta). Potravu šatovníkům poskytují i některé nepůvodní druhy rostlin, jako je mučenka banánová (Passiflora mollissima).[20] Tato liána byla na Havajské ostrovy introdukována, nicméně v některých oblastech představuje dokonce hlavní zdroj potravy šatovníků.[19] Zobák šatovníka šarlatového je zřejmě přizpůsoben konzumaci nektaru místních lobelek. Šatovník jej získává specifickou technikou, při níž se na květu rychle přehoupne hlavou dolů, otočí tělo a zobákem směrem nahoru pronikne do dlouhého, dolů zahnutého květu. Během získávání potravy je značně rychlý a dokáže navštívit desítky květů během několika minut. Jedinci zároveň při hledání kvetoucích stromů podnikají i delší lety, přičemž jde o důležité opylovače místních rostlin.[11][19]
Hnízdění
.jpg.webp)
Šatovník šarlatový je monogamním druhem, nicméně takové chování vykazuje pouze v období hnízdění, zatímco po konci rozmnožovací sezóny spolu páry dále nezůstávají. Pouto mezi členy páru je udržováno skrze časté dvoření, samci se samicím druží zpěvem, pomalým kýváním těla a třepetáním křídly, jakož i přeskakováním z větve na větev nebo krátkými lety, které doplňují zpěvem. Samice zaujímají postavení, během kterého spustí křídla a třepetají s nimi; pokud se přikrčí, je to pro samce signál pokusit se o kopulaci.[21] Vrchol rozmnožování obvykle připadá na období mezi únorem a červnem, kdy také vrcholí období kvetení stromů ohia. Pár si v tuto dobu vydržuje menší území v blízkosti hnízda. Hnízdo samotné je stavěno na vrcholcích stromů ohia.[11] Hnízdo má miskovitý tvar s vnějším průměrem asi 9,5 cm. Je tvořeno především větvičkami a mechy, případně i jinými podobnými materiály, a výstelku tvoří lišejníky, rozdrcené listí a stromová kůra. Na stavbě hnízda se podílí primárně samice, nicméně i samec se zapojuje do konstrukčních prací. Celý proces výstavby hnízda trvá asi 4 až 6 dnů. Asi 1 až 5 dní po dokončení hnízda, obvykle brzy ráno, začíná samice s kladením vajíček. Ta jsou bělavá s různými hnědavými znaky, dosahují délky asi 21 mm. Typická snůška činí dvě vejce. Inkubaci zajišťuje pouze samice, zatímco její partner se v tuto dobu pouze ozývá z blízkých hřadů.[22][23]
.jpg.webp)
Inkubace vajec trvá asi 14 dní.[11] Mláďata jsou nidikolní, rodiče jim tedy musí poskytovat potravu. Ptáčata krmí z většiny samice, nicméně samec zase poskytuje potravu samici. Vylíhnutá mláďata mají zavřené oči, začínají vidět teprve asi za 5 dní.[21] Peří mláďat je především špinavě bílé, částečně i šedavé, hlavně na hlavě. Dospívající ptáci mají peří na těle v zelenavých odstínech, přičemž pera na křídlech a ocasu se pohybují v tmavých barvách.[24] Postupně se mění rovněž tvar zobáku, jenž je zpočátku krátký a rovný, přičemž s přibývajícím časem se prodlužuje a zakřivuje. Mláďata opouštějí hnízdo ve věku asi 3 týdnů. Vyvádí je pouze samice, a sice během noci a v období silných dešťů. Mláďata nicméně i tak pokračují v následování jednoho nebo více rodičů, a to i po několik měsíců. Po skončení hnízdění se šatovníci z rozmnožovacích území rozptylují, přičemž následně mohou tvořit i malá hejna, někdy i mezidruhová. Délka života ve volné přírodě je neznámá.[11][21]
Predátoři, parazité a nemoci
Predátory šatovníků šarlatových jsou káně havajská (Buteo solitarius) a místní poddruh kalouse pustovky (Asio flammeus sandwichensis). Vystaveni jsou však predaci i plenění hnízd od některých nepůvodních druhů, jako je sova pálená (Tyto alba), promyka zlatá (Herpestes auropunctatus), dále též krysy, potkani a kočky.[25] Vážnou hrozbu pro šatovníka šarlatového i ostatní havajské ptactvo však představuje především šíření nepůvodních infekčních onemocnění. Jsou jimi jak ptačí neštovice, jejichž původci jsou velké DNA viry rodu Avipoxvirus, tak ptačí malárie, kterou vyvolávají výtrusovci rodu Plasmodium. Obě onemocnění přenášejí komárovití.[26] Především úmrtnost na ptačí malárii je u tohoto druhu extrémně velká. Studie, během níž bylo experimentálně infikováno deset šatovníků šarlatových, ukázala, že pouhé jedno bodnutí infikovaným komárem stačilo na zabití 90 % zkoumaných ptáků. Přežil pouze jediný samec, jenž se následně stal vůči dalším infekcím imunní. Šatovníci z další skupiny, kteří byli bodnuti několikrát, zemřeli bez výjimky. Samci byli k infekci obecně vnímavější než samice, stejně jako ptáci s nízkou počáteční hmotností ve srovnání s těžšími jedinci. Naproti tomu panenka muškátová (Lonchura punctulata), u níž byla vnímavost k ptačí malárii studována souběžně, přičemž není původním druhem souostroví, byla vůči onemocnění zcela imunní. Výsledky studie demonstrují velký vliv dovlečených onemocnění na endemitní druhy ptactva.[27]
Stav populací
Početnost
_Hosmer_Grove%252C_Haleakala_Nat_Park%252C_Maui_co%252C_Hawaii_-01_(41106135465).jpg.webp)
Souhrnná studie z roku 2013 odhadla celkovou průměrnou velikost všech populací na více než 605 000 jedinců. Počet ptáků je však v rámci hlavního řetězce Havajských ostrovů velmi nerovnoměrně rozprostřen. Největší množství ptáků stále přežívá na hlavním ostrově Havaji, podle odhadů průměrně více než 543 000 jedinců.[28] Populační trendy na hlavním ostrově jsou vcelku smíšené, zatímco na závětrné straně ostrova se zdá být populace silně rostoucí, počet jedinců z návětrné části ostrova převážně klesá (ačkoli severní populace z chráněné oblasti Hakalau se zdají být spíše stabilní).[5][29] Rozporuplné výsledky přináší také odhad početnosti šatovníků v izolované oblasti Kohala. Odhady ze 70. a 80. let hovořily o několika stovkách jedinců, nicméně pozdější studie odhadla průměrnou populaci šatovníků na více než 4 920 ptáků v roce 2017. Rozdílná načasování průzkumů však mohla ovlivnit srovnatelnost výsledků.[5][30]
Na ostrově Maui šatovník šarlatový obývá dvě oblasti na východě a západě ostrova, přičemž tyto populace byly izolovány kvůli postupným ztrátám přirozeného prostředí v důsledku lidských aktivit. Na východě Maui byl ve studii z roku 2013 počet šatovníků odhadnut v průměru na více než 59 800 kusů, přičemž tyto populace opět vykazovaly smíšené trendy: zatímco na severovýchodě Maui mezi roky 1980 a 2011 došlo k jejich úbytku (s předpokládaným poklesem o 34 % během 25 let), jihovýchodní populace se zdály být naopak stabilní až rostoucí (o 22 % během 25 let). Na západním Maui žije pouze malé množství ptáků, přičemž pro nedostatečné množství pozorování nelze jejich početnost odhadnout.[31] Ostrovy Havaj a Maui představují v současnosti hlavní državy šatovníků šarlatových. Na ostrovech Oahu a Molokai jsou počty ptáků pravděpodobně pouze nepatrné.[32] Na ostrově Kauai šatovníci šarlatoví přežívají na náhorní plošině Alakai, přičemž jejich počet zde zřejmě prudce klesá. Studie z roku 2013 odhadla její početnost na průměrně asi 2 550 jedinců, se snížením populací o 92 % za 25 let.[33]
Ohrožení a ochrana

Havajské ostrovy byly výrazně poznamenány nejprve prvotním osídlením Polynésany, později kontaktem se západními kolonizátory z 18. století. Polynéská kolonizace vedla k vyhubení více než 70 známých druhů a poddruhů havajských ptáků, zatímco vliv Evropanů vedl k vymizení dalších více než 20 zástupců místní avifauny. Hrozby představovalo především zavlečení nemocí, ale i některých nepůvodních rostlin a živočichů, jakož i ztráta přirozeného prostředí. Šatovník šarlatový byl prvními Evropany popisován jako jeden z vůbec nejhojnějších ptáků souostroví a ještě na konci 19. století byl podle zpráv evropských přírodovědců široce rozšířen ve všech zalesněných oblastech ostrova, od hladiny moře až po vyšší nadmořské výšky. Nicméně již v první půlce 20. století se začala populace šatovníků šarlatových značně snižovat, ptáci se postupně stahovali do čím dal vyšších nadmořských výšek. Na ostrově Lanai šatovník šarlatový vyhynul.[34]
Hlavním důvodem úbytku šatovníků šarlatových se stalo šíření ptačí malárie, jejímž vektorem je nepůvodní komár Culex quinquefasciatus. Původní ptáci souostroví jsou k podobným infekcím velmi vnímaví. Vzhledem k tomu, že komáři žijí a rozmnožují se především v nižších nadmořských výškách, postupné šíření infekčních onemocnění vedlo k vytlačení šatovníků do výše položených areálů. Ptáci z vyšších nadmořských výšek však stále mohou být malárií ohroženi, a to pokud se přesouvají do nižších stanovišť za potravou. Významnou hrozbu navíc představuje postupující změna klimatu, která může vést k dalšímu rozšiřování malárie do vyšších stanovišť, která jsou zatím z tohoto pohledu chráněna.[5][11][16]
Zvýšení prevalence ptačí malárie v důsledku rostoucích teplot již bylo hlášeno na plošině Alakai i v lesní rezervaci Hakalau na hlavním ostrově. Zatímco mezi lety 1988 až 1992 byla v Hakalau prevalence onemocnění nulová, mezi roky 2001 až 2002 činila 8 %. Zvyšování teplot zároveň vede k tomu, že v oblastech, kde se onemocnění zatím vyskytuje pouze periodicky, může být umožněn jeho stálý přenos.[35] Podle některých odhadů by se území šatovníků šarlatových mezi lety 1990–2010 a 2080–2100 mohlo z důvodu klimatických změn zmenšit o téměř 60 %.[36] Jiná studie modelující dopady globálního oteplování na populace šatovníků šarlatových dokonce předpovídá, že do roku 2100 bude druh na pokraji vyhynutí, přičemž celkový počet párů by měl činit pouze asi 0,2–12,3 % odhadované hustoty před výskytem malárie.[37] Vyjma ptačí malárie je šatovník šarlatový postihován i ptačími neštovicemi, není nicméně jasné, jestli toto onemocnění má vliv na početnost populací.[5]
Na tyto problémy se navíc nabaluje také ztráta stanovišť, stejně jako dopad dalších nepůvodních druhů na přírodu ostrova, přičemž jde o introdukované predátory a některé kopytníky, jejichž vliv narušuje skladbu původního porostu. Specifické postavení mají v tomto ohledu zavlečená prasata, jež svou aktivitou vytvářejí nádrže, v nichž se dále mohou množit komáři. Skrze celé Havajské souostroví se také šíří množství invazních druhů rostlin, a ačkoli se šatovník ostrovní může krmit i těmito druhy, pokles rozmanitosti flóry, jenž je spojen s vlivem invazivních druhů, je obecně problematický, neboť může vést ke snížení dostupnosti nektaru v různých ročních obdobích. Vyjma šatovníků navíc mohou některé epidemie postihovat i rostliny, které jim poskytují potravu. Na začátku 10. let 21. století byla na Havajských ostrovech poprvé zaznamenána houba rodu Ceratocystis, patogen rostlin, který napadá stromy ohia a způsobuje jejich rychlý úhyn. Šíření této houby, jež je opět spojena s teplejšími oblastmi, může také napomoci klimatická změna.[5][35]
Mezinárodní svaz ochrany přírody šatovníka šarlatového k roku 2020 hodnotí jako zranitelný druh a do konce 21. století očekává výrazný pokles jeho populací, s odhadovaným poklesem o 30 – 49 % jen během příštích deseti let.[5] Ochranná opatření se vesměs soustředí na kontrolu infekčních onemocnění a zachování původních stanovišť, tj. především jde o likvidaci nepůvodních druhů a oplocování chráněných stanovišť, aby v nich byl zachován původní ráz ostrova.[11] V roce 2017 byl šatovník šarlatový vyhodnocen Správou Spojených států pro ryby, planě rostoucí rostliny a volně žijící živočichy jako ohrožený druh podle zákona o ohrožených druzích.[38][39] Budoucí možné ochranné snahy mohou zahrnovat i vývoj šatovníků odolných proti malárii pomocí metod genetického inženýrství.[40]
V kultuře
.jpg.webp)
Šatovník šarlatový má významné místo v kultuře a mytologii domorodých Polynésanů. Šatovníci jsou považováni za posvátné a vystupují i v příbězích místních obyvatel. Podle havajských bájí polobůh Maui miloval původní lesní ptáky a daroval jim jasné červené a žluté barvy, přičemž následně umožnil, aby se i ostatní lidé mohli těšit z jejich krásy. Pták iiwi, tedy šatovník šarlatový, měl být zvlášť nádherně vybarven, a navíc se ozýval jedinečným voláním, které rezonovalo po celém lese.[41][42]
Peří šatovníků šarlatových i jiných původních havajských ptáků bylo také vysoce ceněno jako materiál pro výrobu okrasných plášťů a přileb, které náležely vysoce postaveným představitelům ostrova (nejprve pouze mužům, později tyto výsady získaly i ženy). Peří šatovníků bylo pevně vetkáno ve tvaru rozličných ornamentů do jemné síťoviny, jejímž základem byla vlákna kopřivovité rostliny Touchardia latifolia, kterou obyvatelé Havajských ostrovů nazývají olona. Síťovina samotná je pojmenována naepuni a její výroba bývala značně zdlouhavá. Ptáci byli loveni na lep tvořený latexem místních rostlin, jenž byl nanášen na větvičky rozmístěné po lese. Lovci se nazývali kia manu.[42] Šatovník šarlatový někdy sloužil dokonce jako vějička, která měla přilákat jiné žádané druhy ptactva. Domorodí lovci na ostrově Havaji tímto způsobem využívali agresivního chování ptáka druhu moho ozdobný, aby jej mohli ulovit. Moho při snaze napadnout uvězněné šatovníky uvíznul v pasti také.[43] Hotové pláště a přilby mohly obsahovat i více než půl milionu ptačích per. Ptáci, kteří byli lapeni pro okrasná pera, mohli být často uvařeni a snědeni. Navzdory tomu se nezdá, že by lov ze strany domorodců vedl k vymírání místních druhů ptáků, ztráta přirozených stanovišť se z tohoto pohledu ukázala býti větší hrozbou.[24]
Odkazy
Reference
- Červený seznam IUCN ohrožených druhů 2021.3. 9. prosince 2021. Dostupné online. [cit. 2021-12-27]
- JANSEN, Justin J. F. J.; STEINHEIMER, Frank D. The authenticity of ‘I’iwi Drepanis coccinea (G. Forster, 1781) skins from Cook's third voyage: what taxidermy can add to the discussion. Bulletin of the British Ornithologists’ Club. 2017-12, roč. 137, čís. 4, s. 246–260. Dostupné online [cit. 2022-01-06]. ISSN 0007-1595. DOI 10.25226/bboc.v137i4.2017.a2.
- Pratt 2005, s. 269.
- Fancy a Ralph 1998, s. 3.
- BirdLife International. 2020. Drepanis coccinea. The IUCN Red List of Threatened Species 2020: e.T22720844A179228794. https://dx.doi.org/10.2305/IUCN.UK.2020-3.RLTS.T22720844A179228794.en. Accessed on 21 December 2021.
- PIVODA, O. Pozdravy od protinožců. Jména živočichů přejatá z domorodých jazyků Austrálie a Oceánie 2.. ŽIVA AVČR. 2020, čís. 3, s. 158. Dostupné online.
- MUNRO, George Campbell. Birds of Hawaii. New York: Tuttle Publishing 235 s. Dostupné online. ISBN 978-1-4629-0954-4, ISBN 1-4629-0954-X. OCLC 811507080
- KNOWLTON, Jessie L.; FLASPOHLER, David J.; MCINERNEY, N. C. Rotzel. First Record of Hybridization in the Hawaiian Honeycreepers: 'I'iwi (Vestiaria coccinea) × 'Apapane (Himatione sanguinea). The Wilson Journal of Ornithology. 2014-09, roč. 126, čís. 3, s. 562–568. Dostupné online [cit. 2021-12-21]. ISSN 1559-4491. DOI 10.1676/13-054.1.
- LERNER, Heather R.L.; MEYER, Matthias; JAMES, Helen F. Multilocus Resolution of Phylogeny and Timescale in the Extant Adaptive Radiation of Hawaiian Honeycreepers. Current Biology. 2011-11, roč. 21, čís. 21, s. 1838–1844. Dostupné online. DOI 10.1016/j.cub.2011.09.039. (anglicky)
- SMITH, Thomas B.; FREED, Leonard A.; LEPSON, Jaan Kaimanu. Evolutionary Consequences of Extinctions in Populations of a Hawaiian Honeycreeper. Conservation Biology. 1995, roč. 9, čís. 1, s. 107–113. Dostupné online [cit. 2021-12-22]. ISSN 0888-8892.
- Iiwi (Vestiaria coccinea) [online]. Arkive.org [cit. 2021-12-21]. Dostupné online. (anglicky)
- Fancy a Ralph 1998, s. 2.
- Pratt 2005, s. 270.
- ZRZAVÝ, Jan; MIHULKA, Stanislav; STORCH, David; BURDA, Hynek; BEGALL, Sabine. Jak se dělá evoluce : labyrintem evoluční biologie. 4. vyd. Praha: Dokořán, Argo 479 s. ISBN 978-80-7363-763-7, ISBN 80-7363-763-4. OCLC 982100649 S. 73–74.
- Fancy a Ralph 1998, s. 5.
- ʻIʻiwi (Vestiaria coccinea). www.fs.fed.us [online]. [cit. 2021-12-26]. Dostupné online.
- Pratt 2005, s. 271–272.
- PERKINS, R. C. L. Volume I., Part IV. – Vertebrata. In: SHARP, D. Fauna Hawaiiensis or the zoology of the Sandwich (Hawaiian) Islands. Cambridge: Cambridge University Press, 1903. Dostupné online. S. 441–442. (anglicky)
- Fancy a Ralph 1998, s. 4–5.
- Pratt 2005, s. 271.
- Pratt 2005, s. 272.
- Fancy a Ralph 1998, s. 8.
- ‘I’iwi [online]. Kauai Forest Bird Recovery Project [cit. 2022-01-04]. Dostupné online. (anglicky)
- Fancy a Ralph 1998, s. 10.
- Fancy a Ralph 1998, s. 7.
- Fancy a Ralph 1998, s. 9.
- ATKINSON, C. T.; WOODS, K. L.; DUSEK, R. J. Wildlife disease and conservation in Hawaii: Pathogenicity of avian malaria ( Plasmodium relictum ) in experimentally infected Iiwi ( Vestiaria coccinea ). Parasitology. 1995-01, roč. 111, čís. S1, s. S59–S69. Dostupné online [cit. 2022-01-04]. ISSN 0031-1820. DOI 10.1017/S003118200007582X. (anglicky)
- Paxton, Gorresen a Camp 2013, s. 10.
- Paxton, Gorresen a Camp 2013, s. 1, 21–26.
- KEITH, Burnett. Distribution, abundance, and acoustic characteristics of Kohala forest birds. , 2020-06 [cit. 2022-01-05]. Thesis. University of Hawaii at Hilo. . Dostupné online. (anglicky)
- Paxton, Gorresen a Camp 2013, s. 16–18.
- Paxton, Gorresen a Camp 2013, s. 19.
- Paxton, Gorresen a Camp 2013, s. 13–15.
- Paxton, Gorresen a Camp 2013, s. 2.
- ‘I’iwi (Vestiaria coccinea) [online]. Hawaiian Bird Conservation Action Plan, 2012-10 [cit. 2022-01-18]. Dostupné online.
- FORTINI, Lucas B.; VORSINO, Adam E.; AMIDON, Fred A. Large-Scale Range Collapse of Hawaiian Forest Birds under Climate Change and the Need 21st Century Conservation Options. PLOS ONE. 28. 10. 2015, roč. 10, čís. 10, s. e0140389. Dostupné online [cit. 2022-01-08]. ISSN 1932-6203. DOI 10.1371/journal.pone.0140389. PMID 26509270. (anglicky)
- GUILLAUMET, Alban; KUNTZ, Wendy A.; SAMUEL, Michael D. Altitudinal migration and the future of an iconic Hawaiian honeycreeper in response to climate change and management. Ecological Monographs. 2017, roč. 87, čís. 3, s. 410428. IP-071642. Dostupné online [cit. 2022-01-08]. DOI 10.1002/ecm.1253.
- SERVICE, U. S. Fish and Wildlife. ‘I‘iwi Receives Protection under the Endangered Species Act. www.fws.gov [online]. [cit. 2022-01-05]. Dostupné online. (anglicky)
- MCGLASHEN, A. The 'I'iwi, a Besieged Hawaiian Forest Bird, Is Now Listed as Threatened. Audubon [online]. 2017-09-20 [cit. 2022-01-05]. Dostupné online. (anglicky)
- SAMUEL, Michael D.; LIAO, Wei; ATKINSON, Carter T. Facilitated adaptation for conservation – Can gene editing save Hawaii's endangered birds from climate driven avian malaria?. Biological Conservation. 2020-01-01, roč. 241, s. 108390. Dostupné online [cit. 2022-01-05]. ISSN 0006-3207. DOI 10.1016/j.biocon.2019.108390. (anglicky)
- ʻIʻiwi. American Bird Conservancy [online]. [cit. 2022-01-06]. Dostupné online. (anglicky)
- KU‘ULEIALOHA WEAVER, A. ‘I’iwi in Hawaiian Culture [online]. Maui Forest Bird Recovery Project [cit. 2022-01-06]. Dostupné online. (anglicky)
- FULLER, Errol. Extinct Birds. New York: Facts on File, 1987. S. 208–209. (anglicky)
Literatura
- FANCY, S. G.; RALPH, C. J, 1998. Iiwi. In: A. Poole & F. Gill. The Birds of North America, No. 327. Philadelphia: The Birds of North America, Inc.. Dostupné online. (anglicky)
- PAXTON, Eben H.; GORRESEN, P. Marcos; CAMP, Richard J., 2013. Abundance, distribution, and population trends of the iconic Hawaiian Honeycreeper, the ʻIʻiwi (Vestiaria coccinea) throughout the Hawaiian Islands. Reston, VA: U.S. Geological Survey. 63 s. Dostupné online.
- PRATT, H. D, 2005. The Hawaiian Honeycreepers. Oxford: OUP. ISBN 019854653X. (anglicky)
Externí odkazy
 Obrázky, zvuky či videa k tématu šatovník šarlatový na Wikimedia Commons
Obrázky, zvuky či videa k tématu šatovník šarlatový na Wikimedia Commons  Taxon Drepanis coccinea ve Wikidruzích
Taxon Drepanis coccinea ve Wikidruzích