Sympatrická speciace
Sympatrická speciace je proces, jehož prostřednictvím se z jedné, na stejném území žijící populace ancestrálního druhu, vyvíjejí druhy nové. V evoluční biologii a biogeografii jsou "sympatrický" a "sympatrie" pojmy odkazujícími na organismy, jejichž areály výskytu se částečně překrývají nebo jsou dokonce zcela stejné. Tyto organismy tedy alespoň na některých místech žijí společně. Jsou-li tyto organismy úzce příbuzné, tedy jde-li například o sesterské druhy, může být takové rozdělení výsledkem sympatrické speciace. Etymologicky, je slovo sympatrie odvozené z řeckých kořenů συν ("společně", "s") a πατρίς ("vlast" nebo "otčina").[1] S termínem sympatrie přišel v roce 1904 britský evoluční biolog Edward Bagnall Poulton.[2]
Sympatrická speciace je jednou ze čtyř tradičních geografických kategorií fenoménu speciace.[3][4] Nejblíže k ní má speciace parapatrická, při níž dochází k vývoji nových druhů na vzájemně čátečně překrytých územích, dále speciace peripatrická, k níž dochází u sousedních, ale nepřekrývajících se populací, a nakonec speciace alopatrická, u níž jsou populace geograficky zcela odděleny. Tyto kategorie vytváří škálu od nulové (sympatrie) až po úplnou (alopatrie) geografickou segregaci oddělujících se skupin, přičemž v prvním případě neexistuje geografická překážka genetickému křížení skupin, zatímco v posledním případě ke křížení mezi jedinci z obou populací vůbec nedochází.[4]
U mnohobuněčných eukaryotických organismů, považujeme sympatrickou speciaci za neobvyklý, ale možný proces vzniku genetické divergence dceřiných populací vzniklých z jediného parentálního druhu obývajícího stejnou zeměpisnou oblast. Tento proces probíhá za vzniku reprodukční bariéry a může vyústit ve zrod nových druhů. [5] U bakterií může být analogický proces (definovaný jako "původ nových bakteriálních druhů, zaujímajících definovatelné ekologické niky") častější; bakterie jsou totiž méně omezeny homogenizačními vlivy pohlavního rozmnožování a také jsou náchylné k poměrně dramatickým a rychlým genetickým změnám prostřednictvím horizontálního přenosu genů.[6]
Historie
Ernst Mayr, proslulý německý evoluční biolog 20. století, tvrdil, že ke speciaci nemůže docházet bez geografické a tedy i reprodukční izolace.[7] Uvedl, že nevyhnutelným důsledkem sympatrie je genový tok, o němž je známo, že je zodpovědný za stírání genetických rozdílů mezi populacemi. Věřil, že k tomu, aby došlo ke vzniku nových druhů, musí být fyzická bariéra alespoň dočasně přítomna.[8] Tato jeho hypotéza vyvolala v diskusi týkající se sympatrické speciace velkou kontroverzi, původně byla populární a tudíž i poměrně vlivná, dnes už je spíše zpochybňována.[9]
První, kdo navrhl dnes nejrozšířenější hypotézu vzniku sympatrické speciace, byl John Maynard-Smith. Přišel na to, že jsou-li dva ekologické výklenky obsazeny jedním druhem, může vlivem divergující selekce mezi těmito dvěma nikami dojít ke vzniku reprodukční bariéry.[10] Adaptací na co nejlepší biologickou zdatnost v každé z obou ekologických nik mohou z jednoho původního druhu vzniknout dva druhy nové, stále žijící ve stejné oblasti, a to i když se jejich příslušníci navzájem náhodně kříží.[11]
Diskuse
Kvůli téměř nemožnému pozorování sympatrické speciace v reálném čase bylo dočasně velmi obtížné tento jev prokázat.[4] Dlouho se věřilo, a mezi význačné proponenty tohoto názoru patřil i Ernst Mayr, že teorie evoluce přírodním výběrem nemůže vysvětlit, jak by z jednoho druhu mohly vzniknout dva druhy nové, když se poddruhy mohou navzájem křížit.[7] Od Mayrova vrcholného období ve 40. a 50. letech 20. století byly navrženy mechanismy vysvětlující, jak může ke speciaci dojít i za podmínek vzájemného křížení nazývaného též genový tok.[11] V poslední době byly empiricky zkoumány i konkrétní případy sympatrické divergence.[12] Nyní se debata přesouvá spíše k tomu, jak často může k sympatrickým speciacím v přírodě docházet, a kolik procent biodiverzity jí lze vysvětlit.
Obtížnost definice
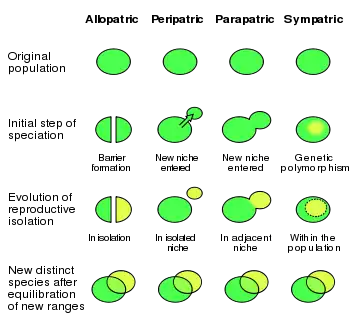
Výzkum možnosti sympatrické speciace vyžaduje zejména ve 21. století, kdy se k výzkumu a predikcím evolučních jevů využívá matematického modelování, řádnou definici tohoto pojmu.[9] Část debat ohledně této formy výběru se točí právě kolem toho, co že to ta sympatrická divergence vlastně je. Použití nejrůznějších definic jednotlivými výzkumníky může vést ke zdržení empirického pokroku v této oblasti. Dichotomie mezi sympatrickou a alopatrickou speciací ji není vědeckou komunitou přijímána, neboť je mnohem užitečnější uvažovat o spojité škále reprodukčních a geografických překryvů mezi druhy. Na jednom konci takového spektra se nachází alopatrie, u níž je mezidruhový překryv nulový (nedochází k žádnému genovému toku), na druhém konci potom leží sympatrie s maximálním genovým tokem a kompletním překryvem areálů.
Různé definice sympatrické speciace spadají obecně do dvou kategorií: definice založené na biogeografii, a definice založené na populační genetice. U čistě geografického pojetí je sympatrická speciace definována jako druh divergující do dvou nástupnických druhů, jejichž areály se zcela překrývají. Tato definice nedostatečně specifikuje původní populaci, takže je nepoužitelná pro modelování.[4]
Definice založené na populační genetice nejsou nutně prostorové či geografické povahy, a někdy mohou být více limitující. Tyto definice pracují s demografickými ukazateli vývoje populace, a to včetně četnosti alel, selekce, velikosti populace, pravděpodobnosti genového toku založené na poměru pohlaví, životních cyklech atd. Hlavním rozdílem mezi oběma typy definic bývá to, nakolik je pro jejich fungování nutná panmixie. Definice sympatrie vycházející z populační genetiky vyžadují náhodné páření, respektive stejnou pravděpodobnost, že se jedinec spáří se zástupcem každého z poddruhů, v jedné či druhé oblasti, a se zástupcem původního nebo nově vzniklého druhu. Tento stav se běžně označuje jako panmixie.[4] To znamená, že populačně-genetické definice vyžadují skutečnou možnost náhodného páření, a ne vždy odpovídají územním definicím toho, co je a co není sympatrie.
Mikro-alopatrií nebo také makro-sympatrií se nazývá situace, kdy se areály výskytu dvou populací sice zcela kryjí, ale ke kontaktu mezi populacemi nedochází, protože obsadily zcela odlišné ekologické niky (například jedna z populací je denní a druhá noční - tomuto se ovšem také někdy říká alochronie). To může být často způsobeno druhově specifickým parasitismem, vytvářejícím mozaikovité rozšíření v krajině (tzv. heteropatrii). Micro-alopatrie je zahrnuta do prostorových definic sympatrie, ale nesplňuje podmínku panmixie, takže podle populačně-genetických definic pod sympatrii nespadá.[4]
Mallet et al. (2002) tvrdí, že nová neprostorová definice neumožňuje vyřešit debatu o tom, zda se sympatrické speciace v přírodě pravidelně vyskytuje či nikoli. Navrhují použít definici prostorovou, ovšem se zahrnutím disperze, neboť toto umožňuje přesněji zachytit možnost genového toku. Podle této publikace by navrhovaná definice měla být použitelná při modelování, a sympatrická speciace se za těchto podmínek zdá být možnou.[8]
Současný stav diskuse
Evoluční teorie i matematické modely předpověděly některé věrohodné mechanismy vedoucí k odštěpení druhů bez nutnosti oddělení populací fyzickou bariérou.[11] Kromě toho existuje několik studií, které objevily speciace probíhající za přítomnosti genového toku (viz sekce důkazy). Molekulární studie ukázaly, že ke speciaci dochází i je-li alopatrie nemožná. Příkladem může být dvojice druhů izolované pouštní palmy. Tyto dva oddělené, ale blízce příbuzné druhy koexistují na stejném ostrově, každý však roste na jiném typu půdy s výrazně odlišným pH.[12] Jde o palmy, jejich pyl je tedy unášen větrem a kdyby již nedošlo ke speciaci zamezující vzniku životaschopných hybridů, mohly by se snadno křížit. Tento příklad je dobrým důkazem možné sympatrické speciace prostřednictvím divergentí selekce v podmínkách konkurence o zdroje, v tomto případě tedy o kus volné půdy.
V současnosti máme k dispozici tento a několik dalších konkrétních příkladů, které však samy o sobě nemohou vypovědět o tom, s jakou frekvencí může za obvyklých podmínek k sympatrické speciaci docházet. Nyní bychom potřebovali objevit rovněž několik dalších případů z méně izolovaných stanovišť. Dosud není známo, za jak velkou část biodiverzity může být zodpovědná právě sympatrická selekce, přičemž podle některých názorů by vzhledem ke zpomalení divergence kvůli panmixii mohl být tento jev sice možný, ale poměrně vzácný (1). Proti tomu stojí řada dalších, kteří považují speciaci bez geografické izolace za možný zdroj velké části pozemské diverzity.[13] Je sice pravdou, že je pro většinu příkladů možné vymyslet těžko vyvratitelné alternativní alopatrické vysvětlení, na druhou stranu s dnešními možnostmi molekulárně-genetickýc postupů bychom snad mohli být schopni teorii sympatrické speciace podpořit.[13]
V roce 2015 byla za použití metod sekvenace DNA pozorována sympatrická speciace u cichlid z malého sopečného kráterového jezera v Africe. Během studie byla objevena složitá směs ekologické separace a pohlavních preferencí vedoucí ke koexistenci a postupnému genetickému oddělení dvou ekomorf, ačkoli stále docházelo k částečné výměně genů.[14][15]
Doklady
K popisu tohoto druhu speciace byla navržena řada modelů. Nejpopulárnějším z nich je model z roku 1966 vytvořený Johnem Maynardem Smithem a odkazující na disruptivní selekci.[16] Maynard Smith navrhl, že by homozygotní jedinci mohli mít za určitých podmínek vyšší biologickou zdatnost nežli jedinci s heterozygotními alelami pro určitý znak. V rámci mechanismu přirozeného výběru, proto může být homozygotnost upřednostňována před heterozygotnostností, což by nakonec mohlo vést ke speciaci. Sympatrická divergence může také vyplývat z pohlavního konfliktu.[17]
Řasy
Zelená mořská řasa Monostroma latissimum vykazuje znaky sympatrické speciace v jihozápadní části japonských ostrovů. Ačkoli jde o panmiktickou populaci, molekulární fylogenetika za použití jaderných intronů odhalila překvapivou diverzifikaci populace.[18]
Rostliny
Jednotlivé události sympatrické speciace jsou mnohem častější u rostlin, které jsou náchylné k zisku a udržení více homologních sad chromozomů, což má za následek stav nazývaný polyploidie. Polyploidní potomci obývají stejné prostředí jako mateřské rostliny (proto jde tedy o sympatrii), jsou ovšem reprodukčně izolované.
Hmyz
Vrtule jabloňová (Rhagoletis pomonella) možná právě s současnosti prochází sympatrickou nebo přesněji řečeno heteropatrickou speciací. Zdá se, že se jablkožravá varianta tohoto druhu, který se původně živil hlohem, odštěpila někdy mezi lety 1800-1850 našeho letopočtu poté, co byla jablka dovezena do Severní Ameriky. Za normálních podmínek už se dnes jablkožravá varianta na hlohu nevyskytuje, a naopak hlohožravá varianta nenapadá jabloně, což je zřejmě první krok na cestě ke vzniku nového druhu.[19][20][21] Prostřednictvím sympatrické speciace se rovněž mohly vyvinout některé druhy parazitických mravenců.[22] Izolované a relativně homogenní stanoviště, jakými jsou například kráterová jezera a ostrovy poskytují nejlepší geografické podmínky k prokázání sympatrické speciace; příkladem může být řada (devět popsaných a desítky nepopsaných) endemických druhů cichlid z nikaragujských jezer, které se vyvinuly sympatricky.[23][24]
Ryby
Několik dokladů sympatrické speciace můžeme najít u afrických cichlid. Ty z oblasti Afrických Velkých jezer vykazují velkou rozmanitost a mnohé studie poukazují na pohlavní výběr jako na možnou příčinu udržování reprodukční izolace. Jedním z nejčastěji studovaných modelů pohlavního výběru u afrických cichlid je samičí volba. Ta se u cichlid vyskytuje v souvislosti s výrazně vyšší maternální investicí do potomstva. U samic dochází k senzorickému biasu ve prospěch nejpodobnějších, případně nejpestřeji zbavených jedinců. [25][26][27] To v jezerech pomáho udržovat sympatrickou speciaci. Cichlidy také při svých námluvách používají akustickou komunikaci. Samečci se během rituálních námluv chvějí a množství pulsů a pulsačních period je signálem pro samičí volbu. Samičí výběr dobrých genů a její senzorický bias patří k rozhodujícím faktorům, přičemž vítězí signály naznačující na příslušnost k danému druhu a na nejlepší biologickou zdatnost, neboť tyto vlastnosti zvyšují šance na přežití potomstva.[28][29] Kompetice mezi samci, která má taktéž vliv na speciace afrických cichlid, je formou intrasexuálního výběru. Rituální boj mezi samci určuje, kteří jedinci budou ve svých reprodukčních snahách nejúspěšnější. To je u sympatrické speciace důležité, neboť druhy s podobnými samci mohou mezi sebou kompetovat o stejné samice. Výhoda v biologické zdatnosti u jednoho druhu může tomuto druhu umožnit invazi do druhu jiného.[30][31] Výzkumy ukazují, že se tento jev vyskytuje především u druhů, které jsou geneticky podobné, a mají tudíž schopnost vzájemného křížení. Zde se potom často objevují nejrůznější fenotypové variace. Další příležitostí pro speciaci je únik do jiných ekologických nik - v jednom jezeru mohou koexistovat různé druhy obživy či hloubka vody, v níž se cichlidy pohybují, mohou napomoci udržení izolace mezi populacemi a později druhy žijícími ve stejném jezeře.
Ptáci
K narušení může dojít rovněž u polygenních vlastností (to jest u znaků řízených spolupůsobením vícero genů). Kupříkladu jedna z tzv. Darwinových pěnkav, pěnkava prostřední (Geospiza fortis) z galapážského ostrova Santa Cruz vykazuje divergenci v té části svého genomu, která odpovídá za morfologii zobáku. Preferovány jsou dvě různé velikosti, zatímco proti nositelům zobáku na půl cesty mezi preferovanými velikostmi působí selekční tlak. Některé vlastnosti někdy nazývané jako magické znaky mohou ovlivňovat speciaci, neboť mají vliv na souboj o zdroje (v tomto případě na získávání potravy) a zároveň mohou hrát roli signálu v rámci pohlavního výběru (různá morfologie zobáku ovlivňuje vokalizaci), což umožňuje vznik vnitrodruhové reprodukční bariéry.[32]
Savci
Srovnatelný jev byl objeven u vrápenců, u nichž se jako magický znak projevuje echolokace. Konstantní frekvenční složka jejich echolkačního signálu určuje velikost lovené kořisti, ale zároveň hraje roli i v sociální komunikaci. Studie provedená na vrápenci filipínském (Rhinolophus philippinensis) ukazuje, že náhlé změny frekvence volání mezi sympatrickými morfami korelují s reprodukční izolací.[33] Mezi dobře prostudované okolnosti vzniku sympatrické speciace patří rovněž situace, kdy se jeden druh hmyzu živí různými rostlinnými druhy. V tomto případě se hmyz postupně specializuje v souboji s odlišnými obrannými mechanismy hostitelských rostlin. (Drès a Mallet, 2002)[34]
Vzácný příklad sympatrické speciace u zvířat je divergence residentních a transientních ekotypů kosatky dravé v severovýchodním Pacifiku.[35] Oba ekotypy obývají stejné vody, ale navzájem se vyhýbají a nekříží se. Tyto dvě formy loví odlišné druhy kořisti, mají odlišní složení potravy, vokalizaci i společenské struktury. Některé rozdíly mezi druhy mohou také vyplývat z odlišností mikrostanovišť. Populace prošly někdy před 200 000 lety hrdlem lahve, při němž se výrazně snižuje velikost populace a tím i genetická diverzita. Následně se mohlo objevit několik rozdílných ekotypů.[36]
Evropský tchoř (Mustela putorius) vykazuje vzácný tmavý fenotyp podobný evropskému norkovi (Mustela lutreola) jehož vzhled je přímo ovlivněn zvláštnostmi lesní potoků.[37]
Alochronie nabízí některé empirické důkazy pro sympatrickou speciaci, neboť mezi alochronickými druhy nacházíme mnoho příkladů nedávno oddělených (sesterských) taxonů.
Odkazy
Reference
V tomto článku byl použit překlad textu z článku Sympatric speciation na anglické Wikipedii.
- Η Πύλη για την ελληνική γλώσσα. www.greek-language.gr [online]. [cit. 2019-03-12]. Dostupné online.
- Poulton, E. B. 1904.
- Futuyma, D. J. 2009.
- Fitzpatrick, B. M.; Fordyce, J. A.; Gavrilets, S. (2008).
- Bolnick, Daniel I.; Fitzpatrick, Benjamin M. (2007).
- King, Stansfield, Mulligan (2006).
- Mayr, Ernst (December 1947).
- Mallet, J.; Meyer, A.; Nosil, P.; Feder, J. L. (2009).
- Jiggins, Chris D. (2006).
- Smith, J. Maynard (1966).
- Kondrashov, Fyodor A.; Kondrashov, Alexey S. (1999).
- Savolainen, Vincent; Anstett, Marie-Charlotte; Lexer, Christian; Hutton, Ian; Clarkson, James J.; Norup, Maria V.; Powell, Martyn P.; Springate, David; Salamin, Nicolas; Baker, William J. (2006).
- Nosil, Patrik (2008).
- "'Darwin's puddle' shows how new species can emerge without geographic separation".
- Malinsky, M.; et al. (2015).
- John Maynard Smith (1966).
- Thierry Lodé La guerre des sexes chez les animaux Eds O Jacob, Paris, 2006
- BAST, F., KUBOTA, S. AND OKUDA, K. 2014.
- McPheron, Bruce A.; Smith, D. Courtney; Berlocher, Stewart H. (1988).
- Smith, D. Courtney (1988).
- Feder, Jeffrey L.; Chilcote, Charles A.; Bush, Guy L. (1988).
- Rabeling, Christian; Schultz, Ted R.; Pierce, Naomi E.; Bacci Jr, Maurício (August 2014).
- Geiger, Matthias F.; McCrary, Jeffrey K.; Schliewen, Ulrich K. (2010).
- Barluenga, Marta; Stölting, Kai N.; Salzburger, Walter; Muschick, Moritz; Meyer, Axel (2006).
- Allender, C.J.; Seehausen, O.; Knight, M.E.; Turner, G.F.; Macleen, N. (2003).
- Egger, B.; Mattersdorfer, K.; Sefc, K.M. (2009).
- Selz, O.M.; Pierotti, M.E.R.; Mann, M.E.; Schmid, C.; Seehausen, O. (2014).
- Amorim, M.C.P.; Simóes, J.M.; Fonseca, P.J.; Turners, G.F. (2008).
- Maruska, K.P.; Ung, U.S.; Fernald, R.D. (2012).
- Seehausen, O.; Schulter, D. (2004).
- Dijkstra, P.D.; Seehausen, O.; Groothuis, T.G.G. (2005).
- Huber, S. K; Leon, L. F. D.; Hendry, A. P; Bermingham, E.; Podos, J. (2007).
- Kingston, Tigga; Rossiter, Stephen J. (2004).
- Begon, Townsend, Harper: Ecology – From individuals to ecosystems, 4th ed., p.10
- Hoelzel, A. R.; Dahlheim, M.; Stern, S. J. (1998).
- "Complete mitochondrial genome phylogeographic analysis of killer whales (Orcinus orca) indicates multiple species".
- Lodé, T. (2001).
Související články
- Polymorfismus (biologie)
- Ekotyp
- Polyploidie
- Adaptivní radiace
- Hybridní speciace
- Kladistika
- Fylogeneze
- Taxonomie
- Wallaceův efekt