Oogonium
Oogonium (pl. oogonia) je označení pro raný (primordiální) oocyt v plodu samičího pohlaví nebo pro samičí gametangium jistých stélkatých rostlin.
Savčí plod
Oogonia jsou formována v plodech samičího pohlaví ve velkém množství mitotickým dělením v raném stadiu fetálního vývoje z primordiálních pohlavních buněk. V lidském těle se začínají vyvíjet mezi 4. a 8. týdnem a v plodu jsou pak přítomny mezi 5. a 30. týdnem.
Struktura
Běžná oogonia v lidských vaječnících jsou kulatá nebo oválná a nacházejí se v sousedících somatických buňkách a oocytech v různých fázích vývoje. Oogonia mohou být rozlišena od sousedících somatických buněk elektronovou mikroskopií při pozorování jejich jádra. Oogoniální jádro obsahuje náhodně rozptýlený fibrilární a granulární materiál, zatímco somatické buňky mají jádro kondenzovanější, mající pod mikroskopem tmavé ohraničení. Oogoniální jádro obsahuje také nápadně hustě uspořádané jadérko. Chromosomy v jádře mitoticky dělícího se oogonia jsou zhuštěná masa obklopená váčky nebo dvojitou membránou.[1]
Cytoplazma oogonií je podobná jako u okolních somatických buněk a také obsahuje velké kulaté mitochondrie s laterálními kristy. Naproti tomu endoplazmatické retikulum je nevyvinuté a skládá se z několika malých váčků. Některé z těchto váčků obsahují cisterny ribozomů a nacházejí se blízko Golgiho aparátu.[1]
Degenerující oogonia jsou pod elektronovým mikroskopem mírně odlišná. Chromozomy se v nich shlukují do nerozlišitelné masy v jádře, mitochondrie a endoplazmatické retikulum se zdají být rozbředlé a narušené. Degenerující oogonia jsou částečně nebo úplně pohlcena přilehlými somatickými buňkami, kde jsou fagocytovány.[1]
Vývoj a diferenciace
V blastocystě savčího embrya se primordiální folikul (pohlavní buňka) vyvine z proximálního epiblastu pod vlivem extraembryonálních signálů. Tyto folikuly poté cestují améboidním pohybem do genitální lišty a poté do nediferencovaných pohlavních orgánů.[2]
Během 4. a 5. týdne vývoje se pohlavní orgány začínají vyvíjet. V nepřítomnosti Y chromozomu se z nich stanou vaječníky. V průběhu diferenciace se vyvíjejí zvláštní vrůsty nazývající se kortikální provazce, v nich se primordiální folikuly shromažďují.[3] [4]
Během 6. až 8. týdne samičího (XX) embryálního vývoje folikuly rostou a diferencují se v oogonia. Oogonia proliferují mitotickým způsobem od 9. do 22. týdne embryonálního vývoje. Do konce 8. týdne se může vyvinout až 600 tisíc oogonií a na konci 5. měsíce je jich až 7 milionů.[3]
Oogonia později buď degenerují, nebo se dále diferencují do primárních oocytů díky asymetrickému dělení. Asymetrické dělení je proces mitózy, při kterém se oogonium rozdělí nerovnoměrně a jedna dceřiná buňka projde následně oogenezí a vznikne z ní oocyt. Druhá dceřiná buňka je identická mateřskému oogoniu. Tento proces proběhne mezi 15. týdnem a 7. měsícem embryonálního vývoje.[2] Většina oogonií se do porodu degeneruje nebo se diferencuje do primárních oocytů.[5][3]
Primární oocyty podstupují oogenezi, ve které nastupuje meióza. Primární oocyty zastaví svůj vývoj v profázi I prvního meiotického dělení a setrvávají v ní až do pohlavního zrání, puberty.[6] Samčí primordiální pohlavní buňky jsou naproti tomu po embryonálním vývoji zastaveny v spermatogoniálním stadiu a nevstupují do spermatogeneze a meiózy za účelem produkce primárních spermatocytů až do pohlavního zrání.[3]
Oogoniální kmenové buňky
Oogonia buď degenerují nebo se diferencují do primárních oocytů, které vstupují do oogeneze a setrvávají v profázi I prvního meiotického dělení až do porodu. Podle této hypotézy se předpokládá, že dospělé savčí samičky postrádají populaci pohlavních buněk, které jsou schopny se obnovovat a regenerovat. Místo toho mají velkou populaci primárních oocytů, které jsou zastaveny v prvním meiotickém dělení až do pohlavního zrání jedince. V pubertě dokončí jeden primární oocyt meiozu a vytvoří vajíčko během každého menstruačního cyklu. Kvůli absenci regenerujících pohlavních buněk a oogonií klesá počet primárních oocytů po každém menstruačním cyklu až do období menopauzy, kdy už v ženském těle žádná populace primárních oocytů není. [2]
Nedávné výzkumy objevily, že obnovy schopná oogonia se mohou nacházet ve výstelce samičích vaječníků primátů, myší i člověka.[7] [2][8] Tyto pohlavní buňky mohou být důležité pro udržování reprodukčních folikulů a vývoj oocytů až do dospělosti. Některé kmenové buňky mohou migrovat z kostní dřeně do vaječníků jako zvláštní zdroj pohlavních buněk. Tyto mitoticky aktivní pohlavní buňky byly identifikovány pomocí označení několika markerů běžných u oocytů. Tyto potenciálně obnovitelné pohlavní buňky byly identifikovány jako pozitivní pro tyto markery esenciálních oocytů.[2]
Stélkaté rostliny
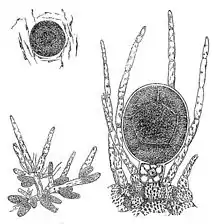
Ve fykologii a mykologii je výraz oogonium označením samičí části gamatangia (samčí je potom antheridium). Oogonium je samičí haploidní gameta v pohlavní fázi rozmnožování určitých druhů „nižších rostlin“, jako jsou např. řasovky (Oomycota).[9][10] Oogonia i antheridia jsou výsledkem meiózy a pohlavní sporulace. Jejich splynutím vznikne diploidní oospora, která později vyklíčí do diploidního somatického stadia životního cyklu stélkatých rostlin.[11]
Struktura
Oogonia některých stélkatých rostlin jsou obvykle kulatá nebo oválná. Jejich obsah je rozdělen do několika uninukleátních (jednojaderných) oosfér. Samčí antheridia jsou naproti tomu oválná a obsahují několik jader. [11] U heterothalických druhů jsou oogonia a antheridia na hyfech rozdílných kolonií. Oogonia těchto druhů mohou být oplodněna jedině antheridii z jiné kolonie a musí být zajištěna nemožnost vlastního oplodnění. Oproti tomu u homothalických druhů jsou antheridia i oogonia vystaveny buď na stejném vláknu hyfy nebo na rozdílných hyfách v rámci jedné kolonie.[11]
Oplodnění
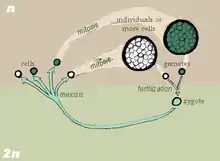
Při běžném oplození u typů thalophyt se antheridia vážnou na oogonia. Antheridia posléze vytvoří oplozovací výběžky spojené s cytoplazmou každé oosféry v oogoniu. Haploidní jádro z antheridií bude poté přesunuto skrz tento výběžek do oosféry a splyne s haploidním jádrem oosféry, vytvoří se diploidní oospora. Oospora je poté připravena vyklíčit a vyrůst do dospělého diploidního somatického stadia vývoje.
Reference
V tomto článku byl použit překlad textu z článku Oogonium na anglické Wikipedii.
- BAKER, T.G., L. L. Franchi. The Fine Structures of Oogonia Oocytes in Human Ovaries. Journal of Cell Science. 1967, s. 213-224. Dostupné v archivu pořízeném dne 2016-03-04. PMID 4933750. (anglicky)
- Germ Stem Cells, A Scientific Summary [online]. New Jersey Medical School [cit. 2012-04-06]. Dostupné v archivu pořízeném dne 2012-01-12. (anglicky)
- JONES, Richard E. Human Reproductive Biology, 2nd Ed.. San Diego: Academic Press, Elsevier, 1997. ISBN 0-12-389775-0. S. 26-40, 90-107, 117-125,. (anglicky)
- CHASSOT, A. A., E.P. Gregory, R. Lavery, M.M. Taketo, D.G. de Rooij, et al. RSPO1/β-Catenin Signaling Pathway Regulates Oogonia Differentiation and Entry into Meiosis in the Mouse Fetal Ovary. PLoS ONE. 2011. Dostupné online [cit. 6 April 2012]. DOI 10.1371. (anglicky)
- Human Emryology, Embryogenesis [online]. [cit. 2012-04-06]. Dostupné online. (anglicky)
- Genetics, Meiosis and Gaetogenesis [online]. www.emich.edu [cit. 2012-04-06]. Dostupné v archivu pořízeném dne 2012-04-30. (anglicky)
- TELFER, Evelyn E., David F. Albertini. The Quest for Human Ovarian Stem Cells. Nature Medicine. 2012, s. 353-354. Dostupné online. DOI 10.1038/nm.2699. (anglicky)
- WHITE, Yvonne A. R., Dori C Woods, Yashushi Takai, OSamu Ishihara, Hiroyuki Seki, Jonathan L. Tilly. Oocyte Formation by Mitotically Active Germ Cells Purified From Ovaries of Reprodutive-Age Women. Nature Medicine. 2012, s. 413-421. Dostupné online [cit. 6 April 2012]. DOI 10.1038/nm.2669. (anglicky)
- Stegenga, H. Bolton, J.J. and Anderson, R.J. 1997. Seaweeds of the South African West Coast. Bolus Herbarium, University of Cape Tow. ISBN 0-7992-1793-X
- Smyth, G.M. 1955. Cryptogamic Botany. vol. 1. McGraw-Hill Book Company
- Sexual Sporulation in Oomycota [online]. [cit. 2012-04-06]. Dostupné v archivu pořízeném dne 2012-04-12. (anglicky)