miRNA
miRNA neboli microRNA jsou jednovláknové řetězce nekódující RNA o délce 21–23 nukleotidů, které se podílejí na regulaci genové exprese. miRNA vznikají transkripcí z genů v DNA, ale následně nedochází k jejich translaci v protein. Namísto toho se každý primární transkript miRNA (tzv. pri-miRNA) páruje s některými vlastními komplementárními bázemi a nakonec se mění na plně funkční miRNA. Tyto molekuly jsou částečně komplementární k určitým molekulám mRNA vyskytujícím se v buňce a jsou schopné regulovat (konkrétně snižovat) tímto výrobu proteinů, které tyto mRNA kódují. K objevu miRNA došlo v roce 1993 v týmu Leeho a jeho spolupracovníků v laboratoři Victora Ambrose,[1] ale termín microRNA je mnohem mladší – pochází z roku 2001 z článků v Science.[2]

MiRNA se vyskytují zejména u rostlin a živočichů, ale vyvinula se u těchto skupin asi nezávisle.[3]
Vznik a úpravy miRNA
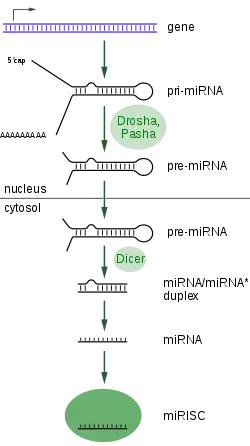
Geny kódující miRNA jsou mnohem delší (obsahují více nukleotidů), než finální upravené miRNA. miRNA je totiž nejprve v jádře hrubě přepsána polymerázami do podoby asi 70 nukleotidů dlouhého řetězce pri-miRNA s čepičkou na 5' konci a poly-A koncem na straně druhé. První úpravy obstarává u živočichů proteinový komplex známý jako Microprocessor complex. Ten je složený z nukleázy jménem Drosha a proteinu Pasha, schopného vázat na sebe dvouvláknovou RNA. Tento komplex mění pri-miRNA na tzv. pre-miRNA[4] Následně pre-miRNA vstupuje do cytoplazmy, kde interaguje s endonukleázou jménem Dicer za vzniku miRNA, jenž se váže do komplexu RISC (RNA-induced silencing complex).[5] Právě RISC je schopen utlumovat expresi genů, jev známý jako RNA interference. U rostlin je celá kaskáda vzniku miRNA mírně odlišná, což je dáno tím, že u rostlin není přítomen protein Drosha a jeho roli v podstatě zastává Dicer.[6]
Funkce
Funkce miRNA zřejmě spočívá v regulaci genů a jejich exprese. Molekuly miRNA jsou komplementární k části jedné nebo několika konkrétních mRNA. Živočišná miRNA vykazuje komplementaritu obvykle k regionu 3' UTR (část mRNA nekódující proteiny, ale vykonávající některé jiné regulační funkce vztahující se k dané molekule mRNA), zatímco rostlinná miRNA je komplementární ke kódujícím regionům messenger RNA. Když se spárují odpovídající řetězce miRNA a mRNA, je obvykle inhibována translace této mRNA v protein.[7] Někdy je namísto toho usnadněn rozklad molekuly mRNA: v tomto případě zřejmě vznik dvouvláknové RNA navozuje v buňce proces podobný RNA interferenci způsobované siRNA molekulami.[7] MiRNA může zřejmě také zasáhnout DNA, která koresponduje s danou mRNA, na níž se miRNA navázala – v tomto případě fungují miRNA spolu s proteiny. označovanými jako miRNP (microribonuclear proteins).
Role miRNA v medicíně
Špatná funkce či regulace miRNA může způsobit v některých případech vážné choroby. Proto je miRNA v centru pozornosti vědců a její výzkum je velmi žádán. Poprvé byla deregulace miRNA v kancerogenezi spojena se vznikem leukémií a lymfomů.[8] Dnes se hledají léky na bázi miRNA, které by pomáhaly například při onemocněních rakovinné povahy a nemocech kardiovaskulární a nervové soustavy.[9]
Některé studie například zjistily, že pokud jsou myši uměle modifikovány tak, aby produkovaly nadměrné množství proteinu c-myc (který má roli ve vzniku rakoviny), umírají na rakovinu mnohem dříve, pokud navíc jejich těla produkují nadměrné množství miRNA.[10] Jiný výzkum prokázal, že miRNA se podílí na regulaci proteinu E2F1, který má roli v proliferaci buněk. V tomto případě se miRNA váže na mRNA a brání tím translaci.[11] Je také možné na základě měření aktivity několika stovek genů kódujících miRNA u pacientů trpících nádorovým bujením zjistit, o jaký typ rakoviny se jedná a z jaké tkáně rakovina vznikla.[12]
Je zřejmé, že miRNa má značný vliv i na činnost srdce. Exprese genů pro miRNA se u lidí s poruchami srdeční činnosti značně odlišuje od zdravých lidí. Zřejmé je to zejména v případě kardiomyopatie,[13][14][15], ale i v případě embryonálního vývoje srdce, poruch vývoje (hypertrofie) a podobně.[16][17][18][19][20][21]
Nejnovější studie také ukázaly, že miRNA produkované rýží (a dalšími rostlinami) nejenže přežívá trávicí proces živočichů, ale dokonce ovlivňuje svojí činností expresi genů. Například MIR168a interferuje s mRNA, ze které se translatuje receptor pro LDL. Dochází tak k utlumení schopnosti živočichů odstraňovat z plazmy LDL.
Další role miRNA
U rostlin jsou miRNA jedním z mechanismů, které patogeny využívají k překonání obranných mechanismů rostlin a usnadnění jejich kolonizace. Stále více důkazů ale naznačuje, že i prospěšné mikroby využívají miRNA k usnadnění symbiózy. Např. mykorhizní houba Pisolithus microcarpus kóduje miRNA, která vstupuje do rostlinných buněk a stabilizuje symbiotickou interakci.[22]
Literatura
Nanjing University School of Life Sciences News Zhang a kol.: "Exogenous plant MIR168a specifically targets mammalian LDLRAP1: an evidence of cross-kingdom regulation by microRNA" Publishing on Cell Research, September 20, 2011.
Reference
V tomto článku byl použit překlad textu z článku MicroRNA na anglické Wikipedii.
- Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell. December 1993, roč. 75, čís. 5, s. 843–54. Dostupné online. DOI 10.1016/0092-8674(93)90529-Y. PMID 8252621.
- Ruvkun G. Molecular biology. Glimpses of a tiny RNA world. Science (journal). October 2001, roč. 294, čís. 5543, s. 797–9. DOI 10.1126/science.1066315. PMID 11679654.
- Archivovaná kopie. www.hoxfulmonsters.com [online]. [cit. 2009-10-21]. Dostupné v archivu pořízeném dne 2009-04-27.
- Denli AM, Tops BB, Plasterk RH, Ketting RF, Hannon GJ. Processing of primary microRNAs by the Microprocessor complex. Nature. November 2004, roč. 432, čís. 7014, s. 231–5. DOI 10.1038/nature03049. PMID 15531879.
- Bernstein E, Caudy AA, Hammond SM, Hannon GJ. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature. January 2001, roč. 409, čís. 6818, s. 363–6. DOI 10.1038/35053110. PMID 11201747.
- Kurihara Y, Watanabe Y. Arabidopsis micro-RNA biogenesis through Dicer-like 1 protein functions. Proc. Natl. Acad. Sci. U.S.A.. August 2004, roč. 101, čís. 34, s. 12753–8. Dostupné online. DOI 10.1073/pnas.0403115101. PMID 15314213.
- CARTHEW, R. W.; SONTHEIMER, E. J. Origins and Mechanisms of miRNAs and siRNAs. Cell.. 2009, roč. 136, čís. 4, s. 642–55. Dostupné online. ISSN 1097-4172.
- Musilova, K; Mraz, M. MicroRNAs in B-cell lymphomas: how a complex biology gets more complex. Leukemia. Macmillan Publishers Limited, 2015, roč. 29, čís. 5, s. 1004–1017. PMID 25541152.
- Vicki Glaser. Tapping miRNA-Regulated Pathways. Genetic Engineering & Biotechnology News. Mary Ann Liebert, Inc., 2008-03-01. Dostupné online [cit. 2008-05-16].
- He L, Thomson JM, Hemann MT, et al. A microRNA polycistron as a potential human oncogene. Nature. June 2005, roč. 435, čís. 7043, s. 828–33. DOI 10.1038/nature03552. PMID 15944707.
- O'Donnell KA, Wentzel EA, Zeller KI, Dang CV, Mendell JT. c-Myc-regulated microRNAs modulate E2F1 expression. Nature. June 2005, roč. 435, čís. 7043, s. 839–43. DOI 10.1038/nature03677. PMID 15944709.
- Lu J, Getz G, Miska EA, et al. MicroRNA expression profiles classify human cancers. Nature. June 2005, roč. 435, čís. 7043, s. 834–8. DOI 10.1038/nature03702. PMID 15944708.
- Thum T, Galuppo P, Wolf C, et al. MicroRNAs in the human heart: a clue to fetal gene reprogramming in heart failure. Circulation. July 2007, roč. 116, čís. 3, s. 258–67. DOI 10.1161/CIRCULATIONAHA.107.687947. PMID 17606841.
- van Rooij E, Sutherland LB, Liu N, et al. A signature pattern of stress-responsive microRNAs that can evoke cardiac hypertrophy and heart failure. Proc. Natl. Acad. Sci. U.S.A.. November 2006, roč. 103, čís. 48, s. 18255–60. DOI 10.1073/pnas.0608791103. PMID 17108080.
- Tatsuguchi M, Seok HY, Callis TE, et al. Expression of microRNAs is dynamically regulated during cardiomyocyte hypertrophy. J. Mol. Cell. Cardiol.. June 2007, roč. 42, čís. 6, s. 1137–41. DOI 10.1016/j.yjmcc.2007.04.004. PMID 17498736.
- Zhao Y, Ransom JF, Li A, et al. Dysregulation of cardiogenesis, cardiac conduction, and cell cycle in mice lacking miRNA-1-2. Cell. April 2007, roč. 129, čís. 2, s. 303–17. DOI 10.1016/j.cell.2007.03.030. PMID 17397913.
- Zhao Y, Samal E, Srivastava D. Serum response factor regulates a muscle-specific microRNA that targets Hand2 during cardiogenesis. Nature. July 2005, roč. 436, čís. 7048, s. 214–20. DOI 10.1038/nature03817. PMID 15951802.
- Xiao J, Luo X, Lin H, et al. MicroRNA miR-133 represses HERG K+ channel expression contributing to QT prolongation in diabetic hearts. J. Biol. Chem.. April 2007, roč. 282, čís. 17, s. 12363–7. Dostupné v archivu pořízeném dne 2020-07-03. DOI 10.1074/jbc.C700015200. PMID 17344217.
- Yang B, Lin H, Xiao J, et al. The muscle-specific microRNA miR-1 regulates cardiac arrhythmogenic potential by targeting GJA1 and KCNJ2. Nat. Med.. April 2007, roč. 13, čís. 4, s. 486–91. DOI 10.1038/nm1569. PMID 17401374.
- Carè A, Catalucci D, Felicetti F, et al. MicroRNA-133 controls cardiac hypertrophy. Nat. Med.. May 2007, roč. 13, čís. 5, s. 613–8. DOI 10.1038/nm1582. PMID 17468766.
- van Rooij E, Sutherland LB, Qi X, Richardson JA, Hill J, Olson EN. Control of stress-dependent cardiac growth and gene expression by a microRNA. Science (journal). April 2007, roč. 316, čís. 5824, s. 575–9. Dostupné online. DOI 10.1126/science.1139089. PMID 17379774.
- Johanna Wong-Bajracharya et al., The ectomycorrhizal fungus Pisolithus microcarpus encodes a microRNA involved in cross-kingdom gene silencing during symbiosis, PNAS January 18, 2022 119 (3)
Externí odkazy
 Obrázky, zvuky či videa k tématu miRNA na Wikimedia Commons
Obrázky, zvuky či videa k tématu miRNA na Wikimedia Commons