Rekombinácia DNA
Rekombinácia je všeobecný termín, ktorý v molekulárnej biológii zahŕňa ľubovoľnú zmenu usporiadania genetickej informácie v rámci jednej alebo medzi viacerými molekulami DNA. Takéto procesy je možné rozdeliť do troch základných tried. Homológna rekombinácia znamená výmenu genetickej informácie medzi dvoma molekulami DNA, ktoré obsahujú región reťazca, ktorý je sekvenčne úplne identický. Miestne-špecifická rekombinácia (z angl. site-directed recombination) znamená výmenu genetickej informácie len v rámci určitej konkrétnej sekvencie DNA. A nakoniec, transpozícia DNA znamená presun určitej genetickej informácie, ktorá má pre tento proces vďaka svojej sekvencii špecifické predpoklady (tzv. skákajúce gény).
Procesy rekombinácie sú pre organizmy dôležité. Hrajú rolu v opravných systémoch DNA, v špeciálnych prípadoch pri replikácii, pri regulácii expresie niektorých génov, sprostredkujú správnu segregáciu chromozómov počas delenia eukaryotických buniek, udržujú genetickú rozmanitosť a zúčastňujú sa cielených genetických zmien usporiadania DNA pri embryonálnom vývine.[1]
Homológna rekombinácia
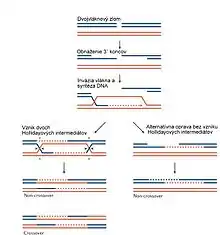
Názov homológna rekombinácia vychádza z toho, že pri rekombinácii majú dve vlákna DNA, ktoré si vymieňajú svoje úseky, sekvenciu s vysokou homológiou, čiže homológnu sekvenciu. Znamená to, že poradie nukleových bází v týchto dvoch vláknach je veľmi podobné.[1]
Homológna rekombinácia u prokaryot
V baktériách hrá rekombinácia DNA rolu predovšetkým pri oprave DNA a raritne sa podieľa na rozširovaní genetickej diverzity pri konjugácii. Rolu hrá často pri replikácii, pri ktorej sa replikačná vidlička zastavuje a prípadne celá rozpadá, ak príde k miestu s chemicky poškodenou DNA. V takom prípade dochádza na poškodenom vlákne k obnaženiu práve nasyntetizovaného vlákna na jeho 3'-OH konci. Následne je toto obnažené vlákno spárované s práve nasyntetizovaným druhým vláknom a pôvodné vlákno s chemicky poškodenou DNA je spárované s druhým pôvodným vláknom. Takto vznikajú v prípade opravy počas replikácie jeden a v prípade opravy mimo procesu replikácie, dva tzv. Hollidayove intermediáty, v ktorom sú dve vlákna DNA navzájom prekrížené. Prekríženie môže byť odstránené buď štiepením vertikálne alebo horizontálne. Pri vertikálnom štiepení sú naštiepené samotné prekrížené vlákna, ktoré sa tak "rozmotajú". Pri horizontálnom štiepení sú štiepené vlákna, ktoré prekrížené nie sú a následne sú obtočené okolo prekrížených vláken tak, aby sa prekrížené vlákna odmotali.[1][2]
Na rekombinácii sa podieľa množstvo enzýmov a proteínových faktorov. Bakteriálna rekombináza sa volá RecA a ide o malý proteín, ktorý sa viaže na DNA a reťazí sa na nej za sebou v smere 5'-3'. Pre činnosť RecA sú potrebné ďalšie proteíny - RecB, RecC, RecD a Pin. K disociácii tohto komplexu dochádza po spárovaní homológnych úsekov DNA v rámci dvoch vláken. Disociácia je sprostredkovaná komplexom RuvC. Chýbajúce fosfodiesterové väzby sú vytvorené DNA ligázou za spotreby ATP. Na konci rekombinácie je vytvorená replikačná vidlička v procese zvanom reštart replikácie nezávislý od počiatku replikácie.[1][2]
Homológna rekombinácia u eukaryot
Podobne ako prokaryoty, používajú eukaryotické bunky homológnu rekombináciu na opravu DNA. Navyše ju však využíva pri meióze, čiže bunkovom delení, pri ktorom vznikajú pohlavné bunky. Pred meiózou sa všetka genetická informácia vo východzích diploidných bunkách zdvojuje a homológne chromozómy k sebe prisadajú práve vďaka rekombinačnému spárovaniu DNA. Počas toho navyše dochádza k homológnej rekombinácii medzi nimi (tzv. crossing over). Táto rekombinácia zvyšuje genetickú variabilitu v potomstve. Aj keď na chromozómoch boli nájdené miesta, v ktorých dochádza k homológnej rekombinácii častejšie, približne platí, že pravdepodobnosť rekombinácie je v celej dĺžke DNA rovnaká. Podobne ako pri oprave DNA, pri homológnej rekombinácii počas meiózy musí dôjsť k dvojvláknovému zlomu DNA na jednom chromozóme a následne k obnaženiu 3'-OH koncov oboch vláken a k ich spárovaniu s homológnymi vláknami na druhom chromozóme. Vznikajú dva Hollidayove intermediáty, ktoré môžu byť odštiepené horizontálne alebo vertikálne. Ak dôjde k homológnej rekombinácii v rámci jednej molekuly DNA, je daný úsek DNA deletovaný a dochádza tak k delécii.[1][2]
Miestne-špecifická rekombinácia
Zatiaľ čo k homológnej rekombinácii môže dôjsť medzi hocijakými homológnymi úsekmi DNA, miestne-špecifická rekombinácia vyžaduje určitú sekvenciu báz DNA. V bunke hrá rolu pri regulácii expresie niektorých génov, pri programovanej zmene usporiadania DNA pri embryonálnom vývoji a je dôležitá pre replikačný cyklus niektorých vírusov.[1] Každá miestne-špecifický rekombinančný systém vychádza z jednej rekombinázy a krátkeho (20 - 200 párov báz) miesta v sekvencii DNA, tzv. miesta rekombinácie. Existujú dva typy takýchto rekombináz a to buď so serínom alebo s tyrozínom v aktívnom mieste. Rekombináza rozpoznáva špecifickú sekvencie na dvoch molekuolách DNA, štiepi ich a kovalentne ich na ich 3' koncoch pripája cez fosfodiesterovú väzbu pomocou -OH skupiny aminokyseliny v aktívnom mieste. Následne jeden 3' koniec spája s voľným 5' koncom druhého vlákna a opačne. Vzniká tak Hollidayov intermediát, v ktorom sú rovnakým spôsobom naštiepené a konvalentne spojené s enzýmom komplementárne vlákna oboch molekúl DNA a následne prepojené tak, aby sa obe novovzniknuté vlákna DNA od seba oddelili. Celý tento proces sa musí zopakovať v ďalších bodoch molekúl DNA a k rekombinácii DNA tak dochádza medzi týmito bodmi.[1] K takejto rekombinácii môže dôjsť aj v rámci jednej molekuly DNA a iba na jednom mieste, v takom prípade dochádza k delécii úseku DNA (k jeho odstráneniu).[1]
Transpozícia
Transpozícia je typ rekombinácie, vďaka ktorému sa po DNA môžu pohybovať tzv. transpozičné elementy alebo transpozony. Transpozony sa do molekuly DNA začleňujú úplne náhodne a môžu tak spôsobiť aj jej smrť. Dá sa povedať, že ide o najjednoduchšie parazitické elementy, ktoré však na druhú stranu mnohokrát pre bunku prinášajú výhodu. Baktérie majú dva základné typy transpozónov - jednoduché transpozóny, ktoré obsahujú len sekvenciu potrebnú pre transpozíciu a gén pre potrebnú transponázu a komplexné transpozóny, ktoré obsahujú navyše ďalšie gény. Tie môžu napríklad kódovať proteín prinášajúci baktériám rezistenciu voči antibiotikám.[1]
Existujú dva druhy transpozície. Pri priamej (jednoduchej) transpozícii je transpozón jednoducho vyštiepený zo svojho pôvodného miesta v DNA a zanecháva za sebou dvojvláknový zlom. Následne je transpozón začlenený do DNA tak, že transponáza predtým štiepi dvojvláknovú DNA na dvoch miestach, vlákna medzi týmito miestami od seba odsadajú a transpozon je vložený medzi takto obnažené vlákna. Časť DNA medzi dvoma štiepnymi miestami je tak zdvojená a po začlenení transpozónu sa nachádza pred ním a za ním. Replikatívna transpozícia sa od jednoduchej odlišuje tým, že je celý transpozón replikovaný a na nové miesto v DNA je začlenená až jeho kópia.[1]
Mechanizmus podobný transpozícii je využívaný pri stavbe širokého spektra protilátok.[1]
Referencie
- NELSON, David L.; COX, Michael M.. Lehninger Principles of Biochemistry. 6. vyd. New York : W. H. Freeman and Company, 2013. ISBN 978-1-4641-0962-1. (anglicky)
- SUNG, P.. Mechanism of homologous recombination: mediators and helicases take on regulatory functions.. Nature reviews. Molecular cell biology, 2006, s. 739-50. (anglicky)