Trepka velká
Trepka velká (Paramecium caudatum) je nálevník běžně se vyskytující v organicky znečištěných vodách po celém světě. Slouží proto jako bioindikátor tohoto znečištění. Trepka velká je při délce 0,17–0,35 mm největší z rodu trepek a je pozorovatelná i pouhým okem.[1] Má asymetrický tvar připomínající botu,[1] který se rozšiřuje směrem k zadní části, což jí přineslo svůj český rodový název. Její buňka je jedna z nejsložitějších v přírodě, [2] a slouží jako modelový organismus nejenom pro výuku biologie na školách, ale i pro výzkum epigenetiky, funkce bičíků nebo procesu endosymbiózy.
 Trepka velká (Paramecium caudatum) | |
| Vědecká klasifikace | |
| Doména | Eukaryota |
| Říše | Chromalveolata |
| Nadkmen | Alveolata |
| Kmen | nálevníci (Ciliophora) |
| Třída | chudoblanní (Oligohymenophorea) |
| Řád | Peniculida |
| Čeleď | Parameciidae |
| Rod | trepka (Paramecium) |
| Binomické jméno | |
| Paramecium caudatum Ehrenberg, 1838 | |
| Některá data mohou pocházet z datové položky. | |
Stavba těla
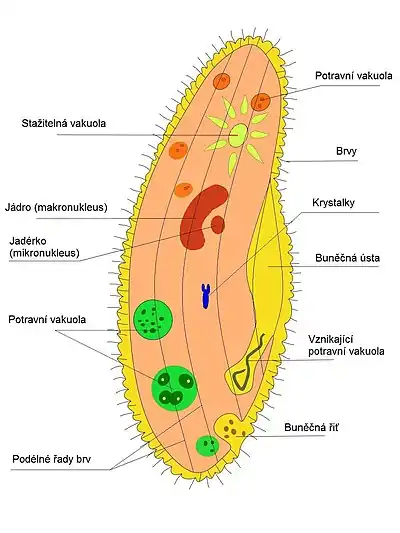
Tělo trepky velké je kryto pelikulou tvořenou trojitou membránou uspořádanou do šestiúhelníkových struktur, z jejichž středů vychází brvy (což jsou v podstatě krátké bičíky[3]), které jí pomáhají k pohybu. Brvy jsou uspořádány do podélných řad, a s výjimkou několika delších na předním konci trepky mají všechny 10-12 μm. Cytoplazma se rozděluje na dvě části, na vnější hustší ektoplazmu obsahující bazální tělíska a trichocysty, a vnitřní polotekutou endoplazmu obsahující ostatní buněčné organely.
Přímo pod pelikulou jsou ukotveny brvy pomocí bazálních tělísek (neboli kinetosomů), které jsou schopny se dělit a vytvářet tak nové brvy. Samotná bazální tělíska vznikají z centriol, které jinak řídí jaderné dělení. Bazální tělíska jsou navzájem propojena pomocí vláken (tzv. kinetodesmální vlákna), které vytváří vnitřní oporu pro brvy.
Trichocysty jsou sekreční organely, které jsou naplněny směsí polypeptidů a malých proteinů uskladněných v podobě tekutého krystalu, přičemž tyto proteiny kóduje až 100 různých genů.[4] Trichocystů je v trepce několik tisíc, jsou tvořeny váčkem o délce asi 4 μm, který je ukončen bodcem. Po aktivaci se trichocysty během milisekund asi 10× prodlouží a uvolní obsah svého váčku. Funkce trichocystů není známa, předpokládá se, že buď slouží k obraně trepky a obsahuje buněčné jedy, nebo slouží k přichycení lovených bakterií.[1]
Buňka trepky obsahuje dvě jádra,[pozn. 1] makronucleus a mikronucleus, projevuje se u ní tedy jaderný dualismus. Větší, ledvinovitě tvarovaný makronucleus zajišťuje vegetativní funkce, je polyploidní, obsahuje tedy několik násobků genetické informace a nachází se v něm několik jadérek. Mikronucleus je menší, kulovitého tvaru a leží v záhybu jadérka, zajišťuje generativní funkce, jadérko neobsahuje. Mikronukleus nebo makronukleos je často infikován symbiotickou bakterií z rodu Holospora, výběr jádra záleží na druhu bakterie. I když silná infekce těmito bakteriemi dokáže trepku zabít, slabá infekce jí zajišťuje odolnost proti řadě stresových podmínek a umožňuje jí přežívat i v brakické vodě (částečně slané).[5]
Na obou koncích trepky se nachází stažitelné vakuoly paprsčitě obklopené kanálky ústícími na povrch buňky. Tato vakuola se s pomocí myofibril stahuje a zbavuje tak trepku přebytku vody, čímž zajišťuje osmoregulaci. Potrava je přijímána buněčnými ústy, kolem kterých jsou čtyři řady brv přihánějících potravu. Dále je potrava vedena buněčným hltanem (cytopharynx) a dochází k vytvoření potravní vakuoly, která je po určité době vyloučena prostřednictvím buněčné řitě.
Rozmnožování
Trepka se rozmnožuje většinou nepohlavně. Jedinec vyroste na dospělém jedinci a po čase se oddělí. Trepky mají i pohlavní rozmnožování. Nazývá se konjugace. Jedinci splynou buněčnými ústy, malé jádro (mikronucleus) prodělá redukční dělení, velké jádro (makronucleus) se rozpadá. Z malého jádra vznikají dělením čtyři jádra, tři z nich zanikají a čtvrté se haploidně rozdělí na dvě. Jádra v buňkách splynou v synkarion a konjuganti se rozestoupí. Následují tři za sebou jdoucí jaderná dělení. Výsledkem konjugace je vznik osmi trepek.
Pohyb
Díky svému hydrodynamickému tvaru a brvám pokrývajícím celé tělo může trepka velká vyvinout rychlost i víc než 1,5 milimetrů za sekundu. Brvy vykonávají kyvadlovitý pohyb, který se skládá z rychlého a silného pohybu vzad, který pohání trepku vpřed, a z pomalého pohybu zpět, který vrací brvu do původní polohy. Samotný pohyb brv probíhá v metachronním rytmu (tedy podobně jako Mexická vlna), jednotlivé řady jsou vzájemně synchronizované, způsob synchronizace ale není prozkoumaný. Výsledný pohyb trepky není přímý, ale v levotočivé šroubovici. Tento nepřímý pohyb je způsoben jednak tím, že brvy se nepohybují přímo zepředu dozadu, ale zabírají poněkud doprava a brvy poblíž buněčných úst přispívají k rotaci trepky.
Školní laboratorní práce
Pozorování trepek pod mikroskopem je častou součástí školních laboratorních prací z biologie. Trepky se získávají ze senného nebo slaměného nálevu. Ten se připraví z chemicky neznečištěné vody z kaluže a trávy či jiné organické hmoty. Nejvíc trepek je v něm asi tři týdny po přípravě. Pro pozorování je nutné přidat do vzorku pod mikroskopem vatu, aby se trepky nemohly pohybovat příliš rychle. V senném nebo slaměném nálevu je možné najít kromě trepek také jiné nálevníky.
Modelový organismus
I když je jako modelový organismus ze skupiny nálevníků častěji využívána tetrahymena thermophila, na některé studie byla využívána i trepka velká. Jedná se především o studium funkce brv (cilie), jejichž struktura je velmi podobná i u vyšších organismů, lidé například využívají brvy (řasinky) ve své průdušnici a ve vejcovodech,[3] porucha v tvorbě řasinek vede u člověka například k Meckelovu syndromu.[6] U trepky velké byla také objevena role glykosylace v regulaci funkce mikrotubulů (součást cytoskeletu).[7] Kromě studia brv jsou trepky využívány také pro studium epigenetiky, především díky jejich jadernému dualismu a schopnosti zbavovat se své nekódující DNA. Relativně nově popsané uchovávání trepky v tekutém dusíku (kryoprezervace) s použitím dimethylsulfoxidu[8] umožňuje přímo studovat evoluci těchto organismů v laboratoři. V tomto ohledu je zajímavá především jako model endosymbiózy, protože její symbiotická bakterie holospora nedokáže přežít mimo ni a oproti volně žijícím bakteriím ztratila část své genetické informace,[5] tento jev je zajímavější o to, že holospora patří do řádu Rickettsiales, ze které pravděpodobně podobným procesem vznikly mitochondrie.[9]
Odkazy
Poznámky
- Dvě jádra má trepka velká v průběhu většiny svého životního cyklu. Během konjugace se ovšem vyskytuje větší množství jader, z nichž většina zaniká. Jiné druhy trepek mohou stabilně obsahovat jiná množství jader, například P. aurelia má dvě mikrojádra a P. multimicronucleatum celou řadu mikrojader
Reference
- KOTPAL, R.L. Modern Text Book of Zoology: Invertebrates. [s.l.]: Rastogi Publications, 2012 883 s. ISBN 8171339034. Kapitola 12. Paramecium caudatum: The Slipper Animalcule.
- BEALE, Geoffrey.; PREER, John R. Paramecium : genetics and epigenetic. Boca Raton: CRC Press, 2008. ISBN 978-0-415-25785-5.
- VINCENSINI, L.; BLISNICK, T.; BASTIN, P. 1001 model organisms to study cilia and flagella.. Biol Cell. Mar 2011, roč. 103, čís. 3, s. 109–30. DOI 10.1042/BC20100104. PMID 21275904.
- MADEDDU, L.; GAUTIER, MC.; VAYSSIÉ, L., et al. A large multigene family codes for the polypeptides of the crystalline trichocyst matrix in Paramecium.. Mol Biol Cell. Jun 1995, roč. 6, čís. 6, s. 649–59. PMID 7579685.
- FUJISHIMA, M.; KODAMA, Y. Endosymbionts in paramecium.. Eur J Protistol. May 2012, roč. 48, čís. 2, s. 124–37. DOI 10.1016/j.ejop.2011.10.002. PMID 22153895.
- CUI, C.; CHATTERJEE, B.; FRANCIS, D., et al. Disruption of Mks1 localization to the mother centriole causes cilia defects and developmental malformations in Meckel-Gruber syndrome.. Dis Model Mech. Jan 2011, roč. 4, čís. 1, s. 43–56. DOI 10.1242/dmm.006262. PMID 21045211.
- REDEKER, V.; LEVILLIERS, N.; SCHMITTER, JM., et al. Polyglycylation of tubulin: a posttranslational modification in axonemal microtubules.. Science. Dec 1994, roč. 266, čís. 5191, s. 1688–91. PMID 7992051.
- KRENEK, S.; BERENDONK, TU. A long-term conservation tool for cell characteristics: cryopreservation of Paramecium caudatum.. Protist. Aug 2009, roč. 160, čís. 3, s. 355–63. DOI 10.1016/j.protis.2009.03.002. PMID 19427813.
- THRASH, JC.; BOYD, A.; HUGGETT, MJ., et al. Phylogenomic evidence for a common ancestor of mitochondria and the SAR11 clade.. Sci Rep. 2011, roč. 1, s. 13. DOI 10.1038/srep00013. PMID 22355532.