Dvoušroubovice
Dvoušroubovice (dvojitá šroubovice, anglicky double helix) je geometrický útvar, který se skládá ze dvou šroubovic se společnou osou (osou šroubovice) a shodnou vzdáleností od této osy, úhlem stoupání a chiralitou, které mají vzájemně posunutou fázi stoupání (někdy se implicitně předpokládá speciální, symetrický případ, kdy jedna šroubovice je souměrná ke druhé vzhledem k ose šroubovice).

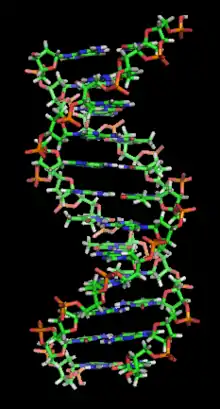
V současnosti je dvoušroubovice nejčastěji spojována s molekulou DNA. Poprvé byla tato biomolekula označena za dvoušroubovici J. D. Watsonem a F. Crickem v roce 1953. Velký význam měla i starší práce Rosalindy Franklin. Tvar dvoušroubovice je velmi stabilní. DNA ho vytváří ze dvou příčin. Molekula, která nese dědičnou informaci, by měla být dvojitá, aby se mohla rozmnožovat (replikovat). Dvoušroubovice je rovněž pevnější než dva samostatné řetězce, protože napínání molekuly do všech stran ji tak snadno nerozdělí.
Tvar dvojité šroubovice má mimoto i mnoho objektů techniky. Příkladem mohou být dvojitá točitá schodiště nebo spojové kabely (například kroucená dvojlinka).
Struktura dvoušroubovice DNA
Základní charakteristika

Dvoušroubovice DNA je tvořena dvěma polynukleotidovými vlákny deoxyribonukleové kyseliny (DNA), které se obtáčí kolem společné osy a interagují spolu. Vlákna jsou tzv. antiparalelní, tzn. směřují opačnými směry[1] – zatímco jedno vlákno můžeme jedním směrem popsat jako 5'-3', druhé je ve stejném směru 3'-5'. Čísla 3' a 5' označují čísla uhlíku na deoxyribóze, na které se upínají fosfátové skupiny v cukr-fosfátové kostře DNA. Dále platí, že jsou vlákna navzájem (alespoň do určité míry) komplementární, tzn. adenin vytváří komplementární pár s thyminem a guanin s cytosinem. Helikální struktura je určena sekundární strukturou, která determinuje konkrétní prostorovou formu stočení (vzdálenost od osy, úhel stoupání, chiralitu a vzájemné posunutí).
Za stabilitu dvoušroubovice DNA jsou zodpovědné především dva faktory: párování bází (vodíkové můstky mezi nukleovými bázemi v rámci páru bází) a dále patrové interakce (stacking interactions) mezi nukleovými bázemi „nad sebou“.[2][3] V obou případech platí, že interakce jsou samy o sobě velmi slabé a jejich síla spočívá v tom, že jich je v dvoušroubovici DNA obrovské množství.[3] Patrové interakce jsou poněkud více závislé na konkrétní sekvenci nukleotidů v jejich oblasti, ale obecně platí, že jsou pro stabilitu dvoušroubovice určující - tedy nikoliv párování bází, nýbrž patrové interakce zodpovídají za stabilitu dvoušroubovice.[2] Roli patrových interakcí popsal český vědec Pavel Hobza.[4][5]
Helikální formy

Většina DNA v buňkách se nachází ve formě tzv. B-DNA, kterou prakticky již v roce 1953 popsali Crick a Watson.[6] Mimo to však byla popsána celá řada jiných (méně či více významných) modelů dvoušroubovice, z nichž některé se zřejmě vyskytují i in vivo, v buňkách živých organismů. Ke známým modelům patří zejména Z-DNA a dále také A-DNA (viz níže). Ukazuje se, že minimálně Z-DNA a H-DNA se mohou v krátkých úsecích polynukleotidových řetězců v genomu živých organismů reálně vyskytovat a zřejmě plní i důležité funkční role.[7] Z-DNA je energeticky velice bohatá a zřejmě se může vyskytovat v místech, kde došlo k silnému negativními nadšroubovicovému vinutí.[6]
| Atribut | A-DNA | B-DNA | Z-DNA |
| Točivá tendence šroubovice (chiralita) | pravotočivá | pravotočivá | levotočivá |
| Opakování | po každém páru | po každém páru | po každých dvou párech |
| Otočení po každém opakování | 32,7° | 34,3° | -60°/2 páry |
| Průměrný počet párů na jedno otočení šroubovice | 11 | 10,5 | 12 |
| Sklon páru k ose | 20° | −1,2° | -9° |
| Vzestup vůči ose na jeden pár | 2,3 Å (0,25 nm) | 3,32 Å (0,34 nm) | 3,8 Å (0,37 nm) |
| Vzestup vůči ose na jednu otočku | 28,2 Å (2,82 nm) | 33,2 Å (3,32 nm) | 45,6 Å (4,56 nm) |
| Průměr | 23 Å (2,3 nm) | 20 Å (2,0 nm) | 18 Å (1,8 nm) |
| Konformace nukleosidu | anti | anti | C: anti, G: syn |
| Konformace ribofuranózy | C3'-endo | C2'-endo | C: C2'-endo, G: C3'-endo |
Každý zdroj však uvádí poněkud odlišná čísla, výše uvedená tabulka tedy není jediná správná. Koneckonců ani B-DNA nacházející se v živých buňkách není ideální Watson-Crickovská struktura. Struktura je zejména silně závislá na sekvenci bází v daném místě.[8]
Žlábky
Důležitou součástí dvoušroubovic jsou žlábky (angl. groove), prohlubňovité struktury nacházející se na dvoušroubovici DNA. Žlábky jsou mimoto přítomny i u dvoušroubovicové RNA, která je však vzácnější.[9] Konkrétní stavba žlábků se liší v závislosti na typu helikálního uspořádání. Žlábky jsou důležitým místem regulace genové exprese, vážou se do nich různé transkripční faktory a podobně. Tzv. velký žlábek (GM)[10] je ten, ve kterém je úhel C(1')–osa šroubovice–C(1') větší než 180°.[10] Ten druhý se označuje jako „malý žlábek“.
Nadšroubovicové vinutí
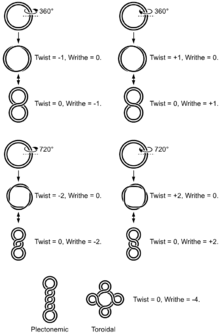
Nadšroubovicové vinutí (supercoiling nebo superhelix[11]) je dodatečné vinutí již existující dvoušroubovice DNA.[12] Jedná se o běžný jev, který však výrazně mění parametry i vlastnosti dvoušroubovice. Podobné struktury bude vytvářet třeba i gumový kroužek, který několikrát převineme přes sebe. Celkový počet vinutí (L) je potom dáno součtem dvoušroubovicového čísla (T, obvykle počet otáček běžné dvoušroubovice B-DNA) a tzv. nadšroubovicového čísla (W). Zmíněné nadšroubovicové číslo slouží jako měřítko superhelicity DNA, jedná se vlastně o počet otáček osy dvoušroubovice kolem osy nadšroubovice.[8] Negativní supercoiling vzniká tehdy, když se nadšroubovice stáčí proti směru hodinových ručiček, tedy naopak, než pravotočivá DNA dvoušroubovice běžné B-DNA. Pozitivní supercoiling má stejnou tendenci jako dvoušroubovice DNA.[13] Na vytváření a potlačování supercoilingu se podílí enzymy známé jako topoisomerázy.
Struktura dvoušroubovice RNA
RNA se obvykle považuje za jednovláknovou molekulu, nicméně to není úplná pravda. I RNA má silnou tendenci k tvorbě kratších či delších dvouvlánkových úseků, jež mají rovněž strukturu dvoušroubovice. Typické je to např. pro některé dsRNA viry. RNA se od DNA liší přítomností jedné z OH skupin ribózy, což má za následek poněkud odlišný druh konformace - jedná se vlastně o A-formu podobnou A-DNA. Také kombinovaný hybridní helix RNA/DNA vytváří A-formu.[14]
Dynamika
Dvoušroubovice není netečná struktura a v čase podléhá poměrně výrazným proměnám. Řada experimentů ukazuje, že dvoušroubovice DNA mohou i za běžné teploty čas od času přechodně denaturovat, tzn. vytvářet lokální „bubliny“, v nichž se rozrušila dvoušroubovice. Toto tzv. dýchání DNA (DNa breathing) je zřejmě do velké míry zodpovědné za nízkou stabilitu krátkých oligonukleotidů.[15] Nejsnáze denaturují oblasti bohaté na AT páry (s nízkým GC obsahem).[16] V buňce mohou dvoušroubovicové číslo (počet obrátek dvoušroubovice) snižovat tzv. DNA helikázy, enzymy, které za spotřeby ATP rozplétají dvouvlákno DNA.[1] Bez helikáz by mimo jiné nebylo možno replikovat a transkribovat DNA – pro správný průběh obou procesů je nutná lokální denaturace. V experimentálních podmínkách se denaturace dvoušroubovice provádí obvykle zvýšením teploty, jde to i chemickou cestou.[16]
Odkazy
Reference
V tomto článku byl použit překlad textu z článku Double helix na anglické Wikipedii.
- Oxford dictionary of biochemistry and molecular biology; revised edition. Příprava vydání R. Cammack et al. New York: Oxford university press, 2006. ISBN 0-19-852917-1.
- YAKOVCHUK, P.; PROTOZANOVA, E.; FRANK-KAMENETSKII, M. D. Base-stacking and base-pairing contributions into thermal stability of the DNA double helix. Nucleic Acids Res.. 2006, roč. 34, čís. 2, s. 564–74. Dostupné online. ISSN 1362-4962.
- SINDEN, Richard R. DNA structure and function. [s.l.]: Gulf Professional Publishing, 1994. 398 s. Dostupné online.
- DNA je nádherný systém [online]. Dostupné online.
- http://www.veda.cz/article.do?articleId=14491
- Albino Bacolla; Robert D. Wells. DNA Secondary Structure. In: Lennarz,W.J., Lane, M.D. Encyclopedia of Biological Chemistry , Four-Volume Set, 1-4. [s.l.]: [s.n.]
- Vladimir N. Potaman; Richard R. Sinden. DNA: Alternative Conformations and Biology. [s.l.]: Landes Bioscience, Madame Curie Bioscience Database, 2000-. Dostupné online.
- VOET, Donald; VOET, Judith. Biochemie. 1.. vyd. Praha: Victoria Publishing, 1995. ISBN 80-85605-44-9.
- BEVILACQUA, P. C.; CECH, T. R. Minor-groove recognition of double-stranded RNA by the double-stranded RNA-binding domain from the RNA-activated protein kinase PKR. Biochemistry. 1996, roč. 35, čís. 31, s. 9983–94. Dostupné online. ISSN 0006-2960.
- KING, Robert C.; STANSFIELD, William D.; MULLIGAN, Pamela K. A Dictionary of Genetics, Seventh Edition. [s.l.]: Oxford University Press, 2006.
- Historie molekulární biologie [online]. Jihočeská univerzita. Dostupné online.[nedostupný zdroj]
- ALBERTS, Bruce , et al. The Molecular Biology of the Cell. [s.l.]: Garland Science, 2002. (4th. ed). Dostupné online. ISBN 0-8153-3218-1.
- Robert K. Murray; Daryl K. Granner; Joe C. Davis; Peter A. Mayes; Victor W. Rodwell. Harper’s Illustrated Biochemistry; twenty-sixth edition. [s.l.]: [s.n.], 2003. ISBN 0-07-138901-6.
- http://cnx.org/content/m11065/latest/
- LEE, O. C.; JEON, J. H.; SUNG, W. How double-stranded DNA breathing enhances its flexibility and instability on short length scales. Phys Rev E Stat Nonlin Soft Matter Phys.. 2010, roč. 81, čís. 2 Pt 1, s. 021906. Dostupné online. ISSN 1550-2376.
- USSERY, D. W. DNA Denaturation [online]. 2001 [cit. 2011-06-14]. Dostupné v archivu pořízeném dne 2010-03-31.
Související články
Externí odkazy
 Obrázky, zvuky či videa k tématu dvoušroubovice na Wikimedia Commons
Obrázky, zvuky či videa k tématu dvoušroubovice na Wikimedia Commons