Virus tabákové mozaiky
Virus tabákové mozaiky je původce nakažlivé choroby tabáku, případně dalších rostlin čeledi lilkovitých. Je významný především tím, že se jedná o první objevený virus na světě vůbec. Virus objevil v roce 1892 ruský vědec Dmitrij Ivanovskij. Genom viru obsahuje RNA a je tvořen pouze několika geny. Virus je schopen infikovat cca 200 druhů rostlin, z toho většina je hospodářsky významných.
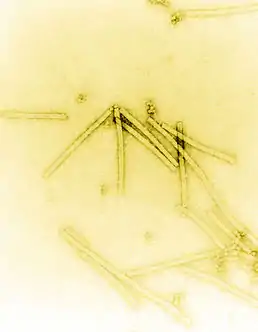 Obrázek viru tabákové mozaiky získaný elektronovým mikroskopem. Zvětšení 160000x. | |
| Baltimorova klasifikace virů | |
| Skupina | IV (ssRNA viry s pozitivní polaritou) |
| Vědecká klasifikace | |
| Čeleď | Virgaviridae |
| Rod | Tobamovirus |
| Druh | Virus tabákové mozaiky (TMV) |
| Některá data mohou pocházet z datové položky. | |
Genetika
Virus tabákové mozaiky (Tobacco mosaic virus) je (+)ssRNA virus o velikosti cca 6 400 nt. 5' konec RNA je opatřen methylguanosinovou čepičkou. Genom kóduje informaci pro 4 proteiny. Dva proteiny tvoří enzym RNA polymerázu, další movement protein a plášťový protein. První otevřený čtecí rámec končí nukleotidem 3417, druhý zahrnuje ten první a prodlužuje ho na 4914 nt, přičemž se překrývá se třetím o 5 nt, a ten končí pouhé 2 nt před začátkem čtvrtého čtecího rámce.[1] Teoretických otevřených čtecích rámců je přítomno více, ale příslušné transláty nebyly v živé tkáni detekovány.
Virus tabákové mozaiky vytváří pomocí hostitelských ribozomů dvě podjednotky specifické virové RNA polymerázy (replikázy) o molekulové hmotnosti 126 a 183 kDa. Translatovaný úsek pro menší replikázu obsahuje stop kodon UAG, který je častokrát přeskočen díky specifické 3' sekvenci CAA-UUA (obecně CAR-YYA), čímž vznikne protein větší, přestože pouze v poměru 20:1 (126:183 kDa).[2] Pro přeskočení stop kodonu jsou i jiné teorie, např. využití hostitelské izoakceptorové tRNA pro tyrosin (antikodon AΨG).
Replikázy vykazují několik enzymatických aktivit - polymerázovou, helikázovou, guanylyl-transferázovou. Poslední aktivita je důležitá pro tvorbu čepičky, která zodpovídá za stabilitu virové RNA a případných kratších mRNA z ní vzniklých.
Netranslatované oblasti jsou krátké, ale přítomné: 5' nekódující oblast (68 nt) vykazuje variabilitu v populaci viru tabákové mozaiky, kdežto 3' nekódující oblast (205 nt) je konzervovaná.[1] Tato konzervovaná 3' oblast zodpovídá za vytvoření dvou pseudouzlů a společně s čepičkou a 5' oblastí zodpovídá za regulaci translace RNA.[3] Za stabilitu virové RNA zodpovídá čepička, která brání hostitelským exonukleázám v degradaci RNA.
Reprodukce viru
Virus se není schopen reprodukovat sám o sobě, k jeho reprodukci je nutný reprodukční aparát hostitelského organismu. Po vstupu do buňky je hostitelskou buňkou transportován k endoplazmatickému retikulu,[5] kde je translatována replikáza, která replikuje virovou RNA. Touto translační a replikační aktivitou je zároveň virion zbaven plášťového proteinu.
Replikace virové RNA probíhá z pozitivních (+) i negativních (-) řetězců RNA, ale přítomnost těchto RNA se liší s průběhem infekce.[6] Jelikož translatovat lze pouze z (+)ssRNA, předpokládá se, že (-) slouží k replikaci finální (+)ssRNA či k produkci mRNA jednotlivých kódovaných proteinů. Následně jsou translatovány plášťové a movement proteiny.
Plášťové proteiny se skládají do duté šroubovice a vytvářejí helikální kapsidu, do které se zanoří virová RNA. Vzniklý virion může za pomocí movement proteinů využít hostitelských buněčných spojení (plazmodezmat) a cytoskeletárního systému (aktinu) pro šíření do dalších buněk.[7]
Zajímavost
V 60. letech 20. století napadl virus tabákové mozaiky většinu pěstebních ploch papričky tabasco. V sedmdesátých letech proto musely být vyšlechtěny nové rezistentní odrůdy (Greenleaf Tabasco).
Reference
- GOELET, P.; LOMONOSSOFF, G. P.; BUTLER, P. J. Nucleotide sequence of tobacco mosaic virus RNA. Proceedings of the National Academy of Sciences. 1982-10-01, roč. 79, čís. 19, s. 5818–5822. PMID: 6964389. Dostupné online [cit. 2020-03-16]. ISSN 0027-8424. DOI 10.1073/pnas.79.19.5818. PMID 6964389. (anglicky)
- SKUZESKI, James M.; NICHOLS, Lindy M.; GESTELAND, Raymond F. The signal for a leaky UAG stop codon in several plant viruses includes the two downstream codons. Journal of Molecular Biology. 1991-03, roč. 218, čís. 2, s. 365–373. Dostupné online [cit. 2020-03-16]. DOI 10.1016/0022-2836(91)90718-L. (anglicky)
- LEATHERS, V; TANGUAY, R; KOBAYASHI, M. A phylogenetically conserved sequence within viral 3' untranslated RNA pseudoknots regulates translation.. Molecular and Cellular Biology. 1993-09, roč. 13, čís. 9, s. 5331–5347. Dostupné v archivu pořízeném dne 2020-07-29. ISSN 0270-7306. DOI 10.1128/MCB.13.9.5331. (anglicky)
- PATTANAYEK, R.; STUBBS, G. Structure of the U2 strain of tobacco mosaic virus refined at 3.5 A resolution using X-ray fiber diffraction.. J Mol Biol. Nov 1992, roč. 228, čís. 2, s. 516–28. PMID 1453461.
- MÁS, Paloma; BEACHY, Roger N. Replication of Tobacco Mosaic Virus on Endoplasmic Reticulum and Role of the Cytoskeleton and Virus Movement Protein in Intracellular Distribution of Viral RNA. The Journal of Cell Biology. 1999-11-29, roč. 147, čís. 5, s. 945–958. Dostupné online [cit. 2020-03-16]. ISSN 0021-9525. DOI 10.1083/jcb.147.5.945. PMID 10579716. (anglicky)
- SHAW, John G. Tobacco mosaic virus and the study of early events in virus infections. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences. 1999-03-29, roč. 354, čís. 1383, s. 603–611. Dostupné online [cit. 2020-03-16]. ISSN 0962-8436. DOI 10.1098/rstb.1999.0412. PMID 10212940. (anglicky)
- CHRISTENSEN, Nynne; TILSNER, Jens; BELL, Karen. The 5′ Cap of Tobacco Mosaic Virus (TMV) Is Required for Virion Attachment to the Actin/Endoplasmic Reticulum Network During Early Infection. Traffic. 2009, roč. 10, čís. 5, s. 536–551. Dostupné online [cit. 2020-03-16]. ISSN 1600-0854. DOI 10.1111/j.1600-0854.2009.00889.x. (anglicky)