Ventrální tegmentální oblast
Ventrální tegmentální oblast příp. ventrální tegmentum, zkr. VTA je skupina neuronů umístěných blízko středové linie na spodní části středního mozku ((lat. tegmentum znamená pokrytí). VTA je zdrojem dopaminergních buněk mesokortikolimbického dopaminového systému a dalších dopaminových drah zapletených do cyklů odměny mozku. VTA hraje důležitou roli v řadě procesů, včetně kognice, motivace, orgasmu,[1] a intenzivních emocí týkajících se lásky, jakož i několika psychiatrických poruch. Neurony v projektu VTA se promítají do mnoha oblastí mozku, od prefrontálního kortexu až po kaudální mozkouvý kmen a několik oblastí mezi nimi.
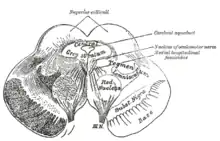
Struktura
Neurobiologové měli velké potíže rozlišovat VTA u lidí a jiných primátů od substantia nigra (SN) obklopující jádra. Původně byla ventrální tegmentální oblast označena také jako „jádro“, ale časem se termín oblast se stal vhodnějším termínem kvůli heterogenním cytoarchitektonickým rysům regionu a nedostatku jasných hranic, které ji oddělují od sousedních oblastí. Vzhledem k selektivní akumulaci vápníku na VTA jsou buňkám VTA označeny jako A10 k odlišení od okolních buněk.
Umístění
Ventrální tegmentální oblast je ve středním mozku mezi několika dalšími hlavními oblastmi. Mammilární tělísko a zadní hypothalamus tvořící diencephalon se táhne rostrálně od VTA. Červené jádro je umístěno laterálně, okulomotorická vlákna ventrálně k VTA. Pons a zadní mozek leží na VTA. Substantia nigra je umístěna laterálně k VTA.
Podjednotky
V roce 1987 Oades identifikoval čtyři primární jádra: nucleus paranigralis (Npn), parabrachialis pigmentosus (Npbp), nucleus interfascicularis (Nif), a nucleus linearis (Nln) caudalis a rostralis. V současné době vědci rozdělují VTA na čtyři podobné zóny, které se nazývají paranigrální jádro (PN), parabrachiální pigmentovaná oblast (PBP), oblast parafasciculus retroflexus (PFR) a rostromediální tegmentální jádro (RMTg), které přibližně dodržuje předchozí rozdělení. Některé definice VTA také zahrnují středová jádra (tj. nucleus interfascicular, rostrální lineární jádro a centrální lineární jádro).
PN a PBP jsou bohaté na dopaminergní buňky, zatímco další dvě oblasti mají nízké hustoty. PFR a RMTg obsahují nízkohustotní tělíska, která jsou pozitivní na buňky obsahující tyrosinhydroxylázu (TH), které mají malou velikost a jsou svěle zabarvena; RMTg je složeno převážně z GABAergních buněk. PN a PBP sestávají převážně ze středně velkých až velkýchTH pozitivních buněčných těl, které se obarví jen mírně.
Vstupy do VTA
Téměř všechny oblasti, které dostávají projekce z projektu VTA dostávají zpětnou vazbu. Ventrální tegmentální oblast je tedy vzájemně spojena se širokou škálou struktur v celém mozku a má úlohu v kontrole funkce ve fylogeneticky novějším a vysoce rozvinutém neokortexu, stejně jako v oblasti fylogeneticky starších limbických oblastí.
VTA není homogenní oblast, protože se skládá z řady neuronů, které se vyznačují různými neurochemickými a neurofyziologickými vlastnostmi. Proto glutaminergní a GABAergní vstupy nejsou výlučně inhibiční ani výlučně excitační. VTA přijímá glutaminergní aferenty z prefrontálního kortexu, pedunculopontinového tegmentálního jádra (PPTg), laterodorálního tegmentálního jádra, subthalamického jádra, jádra lůžka stria terminalis, superior colliculus a laterálních hypotalamických a preoptických oblastí.[2]
GABAergické vstupy do VTA zahrnují nucleus accumbens, ventrální pallidum a rostromediální tegmentální jádro (RMTg). Boční habenula působí inhibicí na dopaminergní neurony ve VTA prostřednictvím ecxitačních neuronů RMTg GABAergic, o kterých se předpokládá, že hrají důležitou roli v chybách predikce odměny.[3]
Excitační glutamatergické afferenty vznikají ze struktur, jež projektují do VTA. Tyto glutamátergické aferenty hrají klíčovou úlohu při regulaci VTA buněk. Když jsou aktivovány glutamátergní neurony, rychlost firingu dopaminových neuronů se zvyšuje ve VTA a indukuje odpálení. Studie ukázaly, že tyto glutamátergické účinky ve VTA jsou kritické pro účinky drog. Naproti tomu konec ventrální tegmentální oblasti (VTA, aka RMTg) promítá do VTA s GABAergic afferenty fungující jako "hlavní brzda" pro VTA dopaminové cesty.[4][5]
Výstupy z VTA
Dvě primární eferentní projekční vláka VTA jsou mezokortikální a mesolimbická dráha, které váží prefrontální kortex a nucleus accumbens.[6][7] Úplný soubor projekcí, z nichž všechny využívají dopamin jako primární neurotransmiter, je uveden níže.[6]
- Projekce ventrální tegmentální oblasti (VTA)[6]
- VTA → Amygdala
- VTA → Cingulate gyrus
- VTA → Hippocampus
- VTA → Nucleus accumbens
- VTA → Čichový bulbus
- VTA → Prefrontální kortex
Vývoj
Vzhledem k tomu, že se vyvíjejí z běžné embryonální tkáně a částečně se překrývají v projekčních polích, nemají dopaminergní buněčné skupiny jasné anatomické hranice. Během vývoje mozku savců, jak substantia nigra tak VTA neurony zpočátku projikují do dorsolaterálního a ventromediálního striata. Při narození však SN dopaminergní neurony promítají výhradně do dorsolaterálního striata a dopaminergní neurony VTA se promítají pouze do ventromediálního striata. K tomuto prořezání spojení dochází prostřednictvím odstranění zbytečných nervových kolaterál.
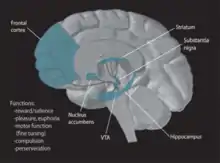
Funkce
VTA slouží v systému odměn, v motivaci, poznávání a drogové závislosti a k několika funkcím a může být zdojem duševních poruch. Bylo také prokázáno, že zpracovává různé typy emocí z amygdaly, kde může také hrát roli při vyhýbání se a podmiňování strachu. Elektrofyziologické záznamy ukázaly, že VTA neurony reagují na nové podněty, neočekávané odměny a smyslově-prediktivní smyslové podněty. Zapojení těchto buněk je konzistentní s kódováním chyby v očekávané odměny.
Dále různé emocionální stavy intenzivní lásky korelovaly s aktivitou ve VTA, což může pomoci vysvětlit obsedantní chování odmítnutých partnerů, protože to souvisí se systémem odměny. Chování hnízda je spojeno se zvýšenou expresí V1aR ve VTA nově spárovaných zebřiček.[8] Exprese V1aR však nesouvisila s mírou samičího bdovu ke zpěvu, což může indikovat selektivní úlohu vazotocinu na VTA a na volbu udržování páru vs. dvoření.[8]
Přítomnost mezerových spojů
VTA má velkou síť GABAergních neuronů, které jsou propojeny přes mezerové spoje. Tato síť umožňuje elektrické vedení, které je podstatně rychlejší než chemické vedení signálů mezi synapsy.[9]
Složení
VTA stejně jako substantia nigra obsahuje melanin zbarvených dopaminergních neuronů.[10] Nedávné studie ukázaly, že dopaminergní neurony zahrnují 50-60% všech neuronů ve VTA[11] což je v rozporu s předchozími důkazy, které zaznamenaly 77% dopaminergní neuronů.[12] Kromě toho existuje značná populace GABAergních neuronů v rostromediálním tegmentálním jádru (RMTg) jako funkčně odlišná struktura mozku.[4][5] Tyto GABAergní neurony regulují zážehy jejich dopaminergních protějšků, které vysílají projekce v celém mozku do následujících oblastí: prefrontální kortex, nucleus accumbens a locus coeruleus. VTA také obsahuje malé procento excitačních glutamátergních neuronů.
Limbická smyčka
Limbická smyčka je velmi podobná motorické smyčce přímé dráhy bazálních ganglií. V obou systémech existují velké excitační vstupy z kortexu do striatum (nucleus accumbens), středový mozek promítá neuromodulační dopaminové neurony do striata, striatum vytváří mezihvězdná spojení s pallidumem a pallidum má výstupy do thalamu, jehož projekty do kortexu, čímž dokončí smyčku. Limbická smyčka je odlišena od motorické smyčky zdrojem a povahou kortikálního vstupu, rozdělení striatum a pallidum, které zpracovávají vstup, zdroj dopaminergních neuronů ze středního mozku a thalamický cíl palidálního výstupu.
Smyčka CA3
Pro hledání odměny je důležité propojení kontextu s odměnou. V roce 2011 skupina výzkumníků zdokumentovala smyčku VTA- CA3, která využívá laterální přepážku jako prostředníka. Jako transsynaptický indikátor použili virus pseudo-vztekliny (PRV) a vstříkli jej do VTA. Zjistili, že jednostranná injekce do VTA má za následek dvoustranné značení PRV na začátku CA3 48 hodin po injekci. Léze caudodorsální laterální septum (cd-LS) před injekcí VTA PRV vedly k významně méně neuronům značeným PRV v CA3. Stimulace CA3 theta vlny vedla ke zvýšeným rychlostem střelby dopaminových buněk ve VTA a ke snížení rychlosti vypalování u neuronů GABA ve VTA. Identita neuronů VTA byla potvrzena neurobiotinovým značením záznamového neuronu a pak histologickým barvením tyrosinhydroxylázy (TH). Dočasná inaktivace CA3 prostřednictvím agonistů GABA zabránila kontextově vyvolanému obnovení páky pro intravenózní kokain.[13]
Autoři navrhli funkční okruhovou smyčku, kde aktivace glutamátergních buněk v CA3 způsobí aktivaci GABAergních buněk v cd-LS, která inhibuje GABA interneurony ve VTA, uvolňuje dopaminové buňky z tonické inhibice a vede ke zvýšené rychlosti střelby pro dopaminových buněk.[13]
Systém odměny
Obvody odměny dopaminu v lidském mozku zahrnují dva projekční systémy od ventrálního středního mozku až po komplex jádra nucleus accumbens- olfactory tubercle. Za prvé, posteromediální VTA a centrální lineární raphe buňky selektivně promítají do ventromediálního striata, které zahrnuje mediální olfaktorální tuberkul a mediální NAC shell. Za druhé, laterální VTA projektuje převážně ventrolaterální striatum, které zahrnuje NAC jádro, mediální NAC shell a laterální olfaktorální tuberkul. Tyto dráhy se nazývají mezo-ventromediální a meso-ventrolaterální striatální dopaminové systémy. Systém mediální projekce je důležitý pro regulaci vzrušení charakterizovaného vlivem a řízením a hraje odlišnou roli v cílovém chování než systém bočních projekcí. Na rozdíl od postranní části se mediální aktivuje nikoli odměnou, ale škodlivými podněty.[14][15] Proto jsou NAC shell a zadní VTA primární oblasti zapojené do systému odměn.
Klinický význam
Poruchy
Dopaminergní neurony substantia nigra a ventrální tegmentální oblast středního mozku vyčnívají do dorsolaterálního caudate / putamen a do ventromediálně lokalizovaného nucleus accumbens, resp. Mezostriatálních a mesolimbických drah. Blízkost těchto dvou cest vede k tomu, že jsou tyto skupiny seskupeny v dopaminergních projekcích. Několik poruch vyplývá z narušení těchto dvou cest: schizofrenie, Parkinsonova choroba a porucha pozornosti s hyperaktivitou (ADHD). Současný výzkum zkoumá jemný rozdíl mezi neurony, které jsou v těchto podmínkách zapojeny, a snaží se najít způsob, jak selektivně léčit specifickou dopaminovou projekci.
Drogová závislost
Primární místa, kde působí návykové léky, jsou nucleus accumbens a ventrální tegmentální oblast. Obvykle jsou považovány za návykové: heroin, kokain, alkohol, opioidy, nikotin, kanabinoidy, amfetamin a jejich analogy. Tyto léky mění neuromodulační vliv dopaminu na zpracování zesilovacích signálů prodloužením účinku dopaminu v nucleus accumbens nebo stimulací aktivace neuronů tam a také ve VTA. Nejčastější zneužívání drog stimuluje uvolňování dopaminu, což vytváří jak jejich odměňování, tak psychomotorické účinky. Kompulzivní chování při užívání drog je výsledkem stálých funkčních změn v mesolimbickém dopaminovém systému vyplývajících z opakované stimulace dopaminu. Molekulární a buněčné adaptace jsou zodpovědné za senzibilizovanou dopaminovou aktivitu ve VTA a podél mesolimbické dopaminové projekce v reakci na zneužívání drog. Ve VTA závislých jedinců se zvyšuje aktivita enzymu tyrosinhydroxylázy syntetizujícího dopamin, stejně jako schopnost těchto neuronů reagovat na excitační vstupy. Posledně uvedený účinek je sekundární v důsledku zvýšení aktivity transkripčního faktoru CREB a zvýšení regulace GluR1, což je důležitá podjednotka receptorů AMPA pro glutamát. Tyto změny v neurálním zpracování by mohly odpovídat za ubývající vliv adaptivních emocionálních signálů při provádění rozhodovacích schopností, protože chování při hledání drog a užívání drog se stává obvyklým a nutkavým.
Experimenty na potkanech ukázaly, že se učí tlačit páku pro podávání stimulačních léků do zadního VTA snadněji než do přední VTA. Další studie ukázaly, že mikroinjekce dopaminergních léčiv do shellu nucleus accumbens zvyšují lokomotorickou aktivitu a průzkumné chování, reakce podmíněného přístupu a předvídavé sexuální chování.
Fenomén abstinenčního stavu nastává proto, že deficit ve funkci odměny iniciuje cyklus nouze, kdy se léky stanou nezbytnými pro obnovení normálního homeostatického stavu. Nedávné výzkumy ukázaly, že i po ukončení závěrečných fází stažení může být chování při hledání léků obnoveno, pokud je vystaveno podnětům souvisejícím s drogami nebo drogami.
Srovnávací anatomie a evoluce
Všechny studie od roku 1964 zdůraznily působivou obecnou podobnost mezi VTA všech savců od hlodavců k lidem. Tyto studie zaměřily své úsilí na krysy, králíky, psy, kočky, vačice, primáty a člověka. Byly pozorovány malé rozdíly, jako jsou změny dorzálního rozsahu buněk A10. Konkrétně, dorzální pík buněk A10 je u primátů ve srovnání s jinými savci rozsáhlejší. Kromě toho se počet dopaminergních buněk ve VTA zvyšuje s fylogenetickou progresí. Například VTA myši obsahuje přibližně 25 000 neuronů, zatímco VTA 33letého muže obsahuje asi 450 000 buněčných těl.[16]
Odkazy
Reference
V tomto článku byl použit překlad textu z článku Ventral tegmental area na anglické Wikipedii.
- HOLSTEGE, G.; GEORGIADIS, J. R.; PAANS, A. M.; MEINERS, L. C.; VAN DER GRAAF, F. H.; REINDERS, A. A. Brain activation during human male ejaculation. The Journal of Neuroscience. 2003, s. 9185–9193. PMID 14534252. (anglicky)
- MORIKAWA, H; PALADINI, CA. Dynamic regulation of midbrain dopamine neuron activity: intrinsic, synaptic, and plasticity mechanisms.. Neuroscience. 15 December 2011, s. 95–111. DOI 10.1016/j.neuroscience.2011.08.023. PMID 21872647. (anglicky)
- WATABE-UCHIDA, Mitsuko; ESHEL, Neir; UCHIDA, Naoshige. Neural Circuitry of Reward Prediction Error. Annual Review of Neuroscience. July 2017, s. 373–394. DOI 10.1146/annurev-neuro-072116-031109. PMID 28441114. (anglicky)
- Bourdy R, Barrot M. A new control center for dopaminergic systems: pulling the VTA by the tail. Trends Neurosci.. November 2012, s. 681–690. DOI 10.1016/j.tins.2012.06.007. PMID 22824232. (anglicky)
- Barrot M, Sesack SR, Georges F, Pistis M, Hong S, Jhou TC. Braking dopamine systems: a new GABA master structure for mesolimbic and nigrostriatal functions. J. Neurosci.. October 2012, s. 14094–14101. DOI 10.1523/JNEUROSCI.3370-12.2012. PMID 23055478. (anglicky)
- [s.l.]: [s.n.] ISBN 9780071481274.
- Nechifor M. Magnesium in drug dependences. Magnes Res. March 2008, s. 5–15. PMID 18557129. (anglicky)
- TOMASZYCKI, ML; RICHARDSON, KK; MANN, KJ. Sex and pairing status explain variations in the activation of nonapeptide receptors in song and motivation regions. Behavioral Neuroscience. October 2016, s. 479–489. DOI 10.1037/bne0000159. PMID 27504854. (anglicky)
- ALLISON, D.W.; OHRAN, A.J.; MAMELI, M.; VALENZUELA, C.F.; SUDWEEKS, S. N.; RAY, A. P.; HENRIKSEN, S. J. Connexin-36 gap junctions mediate electrical coupling between ventral tegmental area GABA neurons. Synapse. 2006, s. 20–31. DOI 10.1002/syn.20272. PMID 16575850. (anglicky)
- http://cogprints.org/1390/2/vta-rev-87.pdf
- MARGOLIS, EB; LOCK, H; HJELMSTAD, GO; FIELDS, HL. The ventral tegmental area revisited: Is there an electrophysiological marker for dopaminergic neurons?. J. Physiol.. 2006b, s. 907–24. DOI 10.1113/jphysiol.2006.117069. PMID 16959856. (anglicky)
- JOHNSON, SW; NORTH, RA. Two types of neurone in the rat ventral tegmental area and their synaptic inputs.. The Journal of Physiology. May 1992, s. 455–68. DOI 10.1113/jphysiol.1992.sp019136. PMID 1331427. (anglicky)
- LUO, Alice; TAHSILI-FAHADAN, P.; WISE, R. A.; LUPICA, C. R.; ASTON-JONES, G. Linking Context with Reward: A Functional Circuit From Hippocampal CA3 to Ventral Tegmental Area. Science. July 2011, s. 353–356. DOI 10.1126/science.1204622. PMID 21764750. Bibcode 2011Sci...333..353L. (anglicky)
- Ikemoto S. Dopamine reward circuitry: two projection systems from the ventral midbrain to the nucleus accumbens-olfactory tubercle complex. Brain Research Reviews. November 2007, s. 27–78. DOI 10.1016/j.brainresrev.2007.05.004. PMID 17574681. (anglicky)
- Brischoux F, Chakraborty S, Brierley DI, Ungless MA. Phasic excitation of dopamine neurons in ventral VTA by noxious stimuli. Proceedings of the National Academy of Sciences of the United States of America. March 2009, s. 4894–4899. DOI 10.1073/pnas.0811507106. PMID 19261850. Bibcode 2009PNAS..106.4894B. (anglicky)
- OADES, R.D; HALLIDAY, G.M. Ventral tegmental (A10) system: Neurobiology. 1. Anatomy and connectivity. Brain Research Reviews. 1987, s. 117–165. DOI 10.1016/0165-0173(87)90011-7. (anglicky)
Související články
Externí odkazy
 Obrázky, zvuky či videa k tématu Ventrální tegmentální oblast na Wikimedia Commons
Obrázky, zvuky či videa k tématu Ventrální tegmentální oblast na Wikimedia Commons