Pomocný T lymfocyt
Pomocné T lymfocyty (Th lymfocyty, Th buňky), známé také jako CD4 buňky, jsou typem T lymfocytů, které mají důležitou roli v imunitním systému, konkrétně v adaptivním imunitním systému. Th lymfocyty uvolňují cytokiny, kterými aktivují ostatní imunitní buňky, a dále pomáhají regulovat imunitní odpověď. Jsou nezbytné pro izotypový přesmyk u B lymfocytů, pro aktivaci a růst cytotoxických T lymfocytů, a aktivaci fagocytů, např. makrofágy.
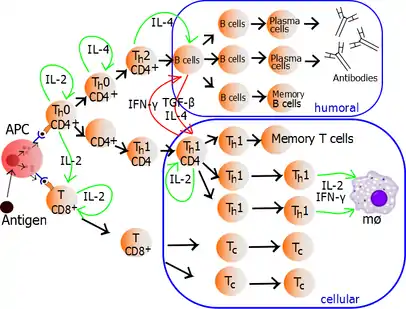
Zralé Th lymfocyty exprimují na svém povrchu protein CD4, proto jsou označovány jako CD4+ T buňky. CD4+ buňky jsou obecně brány jako pomocné T buňky imunitního systému. Například, když antigen prezentující buňka (APC) vystavuje antigen prostřednictvím MHC II. třídy, CD4+ lymfocyt pomůže těmto buňkám prostřednictvím kombinace buněčných interakcí (CD40 ligand) a cytokinů.
Aktivace nezkušených pomocných T lymfocytů
Po vývoji T lymfocytů v thymu se tyto buňky, nazývané recent thymic emigrants (RTE), dostávají z thymu a směřují do sekundárních lymfatických orgánů (slezina a lymfatické uzliny). Následné vyzrávání ve zralé nezkušené T lymfocyty (nezkušené T lymfocyty - buňky, které se doposud nesetkaly se svým specifickým antigenem), je spojeno se sníženou expresí povrchových markerů souvisejících s RTE, např. CD31, PTK7, komplementové receptory 1 a 2 (CR1, CR2), a produkcí interleukinu 8.[1][2] Stejně jako ostatní T lymfocyty i nezkušené T lymfocyty exprimují komplex T buněčného receptoru (TCR) a molekuly CD3. TCR se skládá z konstantních a variabilních oblastí, variabilní oblast rozpoznává antigen. CD4+ T buňky rozpoznávají antigen prezentovaný v komplexu MHC II. třídy, který se obecně vyskytuje pouze na povrchu profesionálních antigen prezentujících buněk (APCs). Mezi profesionální APC řadíme dendritické buňky, makrofágy a B lymfocyty, ale pouze dendritické buňky jsou schopny exprimovat MHC II. třídy konstitutivně.
Rozpoznávání (signál 1)

V průběhu imunitní odpovědi, profesionální antigen prezentující buňky (APCs) pohlcují cizí materiál (obvykle bakterie nebo viry), zpracují jej a poté vycestují z místa infekce do lymfatických uzlin. V lymfatických uzlinách začnou APCs prezentovat antigenní peptidy vázané v komplexu MHC II. třídy, to vede k aktivaci CD4+ T lymfocytů, které exprimují specifické TCR proti prezentovanému peptidu.
Když Th lymfocyt rozpozná antigen na APC, komplex TCR-CD3 (na T lymfocytu) se silně váže na komplex antigenní peptid-MHC II. třídy (na APC). Molekula CD4, která je koreceptorem komplexu TCR, se váže na jiný úsek molekuly MHC II. třídy. Tyto interakce přivádějí zúčastněné proteiny blíže k sobě, což umožňuje intracelulárním kinázám proteinů TCR, CD3 a CD4 se vzájemně aktivovat fosforylací. Pomocí fosfatázy přítomné na intracelulárním úseku molekuly CD45, tyto molekuly aktivují hlavní intracelulární signalizační dráhu Th lymfocytů. Tato signalizační dráha se označuje jako Signál 1 aktivace T lymfocytů, jelikož se jedná o první a primární proaktivační signál pomocných T lymfocytů. Při následném setkání se stejným antigenem dochází právě touto cestou k aktivaci paměťových T lymfocytů.
Vazba komplexu antigenní peptid-MHC s TCR a molekulou CD4 může také pomoci adhezi APC a Th lymfocytů, během aktivace Th. Primárně se však adheze v této buněčné interakci účastní molekuly LFA-1 (Th lymfocyty) a ICAM (APC).
Ověření (signál 2)
Po obdržení prvního signálu musí být nezkušený T lymfocyt aktivován druhou nezávislou biochemickou signální dráhou, známou jako Signál 2. Tento ověřovací krok je ochranným opatřením, které zajišťuje, že T lymfocyt reaguje na cizí antigen. Při nepřítomnosti Signálu 2 během rozpoznání antigenu T lymfocyt předpokládá, že se jedná o autoreaktivní reakci a dochází k anergii T lymfocytu. Anergní T lymfocyt není schopný reagovat na žádný antigen v budoucnu, a to ani v případě, že budou přítomny oba signály. Obecně se věří, že anergní buňky cirkulují po celém těle, dokud neprojdou apoptózou.
Druhý signál zahrnuje interakci mezi CD28 na CD4+ T lymfocytech a proteiny CD80 (B7.1) nebo CD86 (B7.2) na profesionálních APCs. Jak CD80, tak CD86 aktivují receptor CD28. Všechny uvedené proteiny se nazývají kostimulační molekuly.
Ačkoli je ověřovací fáze nezbytná pro aktivaci nezkušených pomocných T lymfocytů, její důležitost je možné demonstrovat na podobné aktivaci CD8+ cytotoxcických T lymfocytů. Nezkušené CD8+ T lymfocyty spoléhají na aktivaci kostimulační molekuly CD28, která tímto způsobem potvrzuje cizí původ rozpoznaného antigenu (CD80/CD86 jsou exprimovány aktivními APCs). Molekula CD28 hraje významnou roli při snížení rizika autoimunitních reakcí T lymfocytů proti hostitelským antigenům.
Když jsou obě signální dráhy spuštěny, dochází k aktivaci doposud nezkušeného T lymfocytu. Druhý signál je poté již zbytečný a pro další aktivaci buňky je zapotřebí pouze Signál 1. To se týká především paměťových T lymfocytů, které jsou součástí adaptivní imunity. Při reinfekci stejným patogenem dochází k rychlejší imunitní odpovědi právě díky tomu, že T lymfocyty již prodělali ověření (Signál 2).
Proliferace
Jakmile je dokončena aktivace oběma signály, pomocný T lymfocyt je schopen proliferovat. Toho je dosaženo pomocí silného růstového faktoru T lymfocytů, zvaného interleukin 2 (IL-2), který je sekretován autokrinně. Aktivované T lymfocyty také produkují alfa podjednotku receptoru pro IL-2 (CD25 nebo IL-2R), což umožňuje expresi funkčního receptoru, který po vazbě IL-2, aktivuje proliferační dráhu T lymfocytů.
Autokrinně nebo parakrinně sekretovaný IL-2 se váže na stejnou Th buňku nebo na sousední Th buňky prostřednictvím IL-2R, to vede k proliferaci a klonální expanzi T lymfocytů. Pomocné T lymfocyty, které obdržely aktivační signály a proliferují, se stávají Th0 lymfocyty, které sekretují IL-2, IL-4 a interferon gama (IFN-γ). Th0 lymfocyty se v závislosti na přítomnost cytokinů diferencují v Th1 nebo Th2 lymfocyty. Prostředí IFN-γ vede k produkci Th1 buněk, zatímco prostředí IL-10 a IL-4 tvorbu Th1 inhibuje. Naproti tomu prostředí IL-4 vede k produkci Th2 buněk a prostředí IFN-γ tvorbu Th2 inhibuje. Je důležité poznamenat, že cytokiny jsou pleiotropní a mají vliv na řadu funkcí imunitní odpovědi.
Stanovení funkční odpovědi T lymfocytů
Pomocné T lymfocyty jsou schopny ovlivnit řadu imunitních buněk, T buněčná odpověď (včetně extracelulárních signálů jako jsou cytokiny) je nezbytná pro úspěšné překonání infekce. Aby byl pomocný T lymfocyt efektivní, musí určit které cytokiny zajistí nejúčinnější nebo nejprospěšnější reakci imunitního systému vůči hostiteli. Porozumět jak pomocné T lymfocyty reagují na imunitní výzvy je v současnosti velkým tématem v imunologii, tato znalost může být užitečná pro léčbu onemocnění a zvyšování účinnosti očkování.
Model Th1/Th 2
Proliferující pomocné T lymfocyty se diferencují do dvou hlavních podtypů známých jako Th1 a Th2 lymfocyty (také označované jako pomocné T buňky typu 1 a 2).
Th1 pomocné lymfocyty vedou ke zvýšení buňkami zprostředkované imunitní odpovědi, převážně proti intracelulárním bakteriím a prvokům (protozoa). Jsou spouštěny pomocí IL-12 a jejich efektorové cytokiny jsou IFN-γ a IL-2. Hlavními efektorovými buňkami Th1 imunitní reakce jsou makrofágy, dále pak CD8+ cytotoxické T lymfocyty, IgG B lymfocyty a IFN-γ CD4+ T lymfocyty. Klíčové transkripční faktory Th1 buněk jsou STAT4 a T-bet. Sekretovaný IFN-γ může aktivovat makrofágy k fagocytóze a štěpení intracelulárních bakterií a prvoků. IFN-γ navíc umožní aktivovat iNOS (inducibilní NO syntáza) za vzniku volných radikálů oxidů dusíku, které přímo zabíjí intracelulární patogeny.
Nadměrná aktivace Th1 imunitní odpovědi proti autoantigenům způsobuje Imunopatologickou reakci 4. typu, tzv. hypersenzitivitu opožděného typu (DTH). Diabetes mellitus typu 1 patří do této kategorie autoimunitních chorob.[3]
Th2 pomocné lymfocyty vedou k humorální imunitní odpovědi, převážně proti extracelulárním parazitům a helmintům. Jsou spouštěny pomocí IL-4 a IL-2, jejich efektorové cytokiny jsou IL-4, IL-5, IL-9, IL-10, IL-13 a IL-25. Hlavními efektorovými buňkami Th2 imunitní reakce jsou eozinofily, bazofily, žírné buňky, dále B lymfocyty a IL-4/IL-5 CD4+ T lymfocyty. Klíčové transkripční faktory jsou STAT6 a GATA3.[4] IL-4 stimuluje B lymfocyty k produkci protilátek třídy IgE. IgE protilátky dále stimulují žírné buňky k uvolňování histaminu, serotoninu a leukotrinů, které zvyšují acidifikaci žaludeční tekutiny, což vede k vyloučení helmintů. IL-5 aktivuje eozinofily, které napadají helminty. IL-10 potlačuje funkci dendritických buněk a také diferenciaci do Th1 lymfocytů.
Nadměrná aktivace Th2 imunitní odpovědi proti autoantigenu způsobuje alergii a přecitlivělost. Alergická rýma, atopický ekzém a astma patří do této kategorie autoimunitních chorob.[3] Kromě exprese rozdílných cytokinů, Th2 lymfocyty se liší od Th1 lymfocytů v povrchových znacích, které činí Th2 lymfocyty méně náchylné vůči indukci buněčné smrti.[5][6]
| Typ 1/Th1 | Typ 2/Th2[3] | |
|---|---|---|
| Partnerské buňky | Makrofág, CD8+ T lymfocyt | B lymfocyt, eosinofil, žírná buňka |
| Produkované cytokiny | Interferon-y a TNF-β Produkce IL-2 a IL-10 zaznamenána u aktivovaných Th1 lymfocytů | IL-4, IL-5, IL-6, IL-9, IL-10, IL-13 |
| Imunitní stimulace | Buněčný imunitní systém. Maximalizuje účinnost zabíjení u makrofágů a proliferaci cytotoxických CD8+ T lymfocytů. Podpora produkce IgG (opsonizující protilátky) | Humorální imunitní systém. Stimuluje proliferaci B lymfocytů, indukuje izotypový přesmyk protilátek. Zvyšuje produkci neutralizačních protilátek (IgG, IgM, IgA, IgE). |
| Další funkce | IFN-y zvyšuje produkci IL-12 dendritickými buňkami a makrofágy a skrze pozitivní zpětnou vazbu, IL-12 stimuluje produkci IFN-y v Th lymfocytech a tím podporuje diferenciaci v Th1 lymfocyty. IFN-y také inhibuje produkci IL-4, čímž inhibuje vznik Th2 lymfocytů. | Th2 imunitní odpověď podporuje vlastní profil pomocí dvou různých cytokinů.
IL-4 působí na pomocné T lymfocyty a podporuje tvorbu Th2 skupiny lymfocytů, včetně samotného IL-4. IL-10 inhibuje řadu cytokinů, včetně IL-2 a IFN-y v Th lymfocytech a IL-12 v dendritických buňkách a makrofázích. Kombinace účinků obou cytokinů naznačuje, že jakmile se T lymfocyt diferencuje do Th2 odpovědi, produkce jeho cytokinů udržuje tento stav a dále podporuje vznik Th2 odpovědi v ostatních buňkách. |
Ačkoli známe typy pomocných T lymfocytů a jejich produkované cytokiny, méně chápeme způsob, jaký se rozhoduje o výběru konkrétního typu. Mnoho důkazů naznačuje, že značný vliv má typ APC prezentující antigen T lymfocytu. Další důkazy navrhují vliv koncentrace antigenu během prvotní aktivace T lymfocytu, případně vliv přítomnosti některých cytokinů uvedených výše.
Th17 pomocné lymfocyty
Th17 pomocné lymfocyty jsou typem pomocných T lymfocytů, které se vývojově liší od Th1 a Th2 typů produkcí interleukinu 17 (IL-17). IL-17 je prozánětlivým cytokinem, který napomáhá v boji proti extracelulárním patogenům a houbovým infekcím.
Omezení modelu Th1/Th2
Interakce mezi cytokiny Th1/Th2 modelu může být v případě některých zvířat komplikovanější. Například Th2 cytokin IL-10 inhibuje u člověka produkci cytokinů obou Th typů. Lidský IL-10 potlačuje proliferaci i produkci cytokinů všech T lymfocytů a také aktivitu makrofágů, naopak dále stimuluje plazmatické buňky a zajišťuje tak stálou tvorbu protilátek. Jako takový tedy IL-10 u člověka nepodporuje Th2 imunitní odpověď, ale brání nadměrné stimulaci pomocných T lymfocytů, při současné maximalizaci tvorby protilátek.
Existují další typy T lymfocytů, které mohou ovlivnit expresi a aktivaci pomocných T lymfocytů, například přirozené regulační T lymfocyty (Treg), společně s méně běžným typem Th3 lymfocytů. Termíny "regulační" a "potlačující" se po objevu CD4+ lymfocytů staly nejednoznačné, protože pomocné T lymfocyty jsou schopny regulovat (a potlačovat) svou vlastní imunitní odpověď, ačkoli nejsou typicky regulačními lymfocyty.
Hlavním rozdílem mezi regulačními a efektorovými T lymfocyty je, že regulační buňky obvykle regulují a potlačují imunitní odpověď, zatímco efektorové buňky obvykle začínají s produkcí cytokinů podporujících imunitní odpověď, a následně přecházejí k produkci inhibičních cytokinů. Právě inhibiční cytokiny jsou znakem Th3 lymfocytů, které se po počáteční aktivaci a produkci cytokinů, transformují v regulační typ buněk.
Regulační T lymfocyty, stejně jako Th3 lymfocyty produkují transformační růstový faktor β (TGF-β) a IL-10. Oba cytokiny mají inhibiční funkci vůči pomocným T lymfocytům. TGF-β potlačuje aktivitu většiny imunitního systému.
Charakterizace nového typu T lymfocytů, v podobně Th17 lymfocytů[7], vyvolala další pochybnosti o základní Th1/Th2 modelu. Tyto nové IL-17 produkující buňky byly prvně popisovány jako patogenní populace buněk spojené s autoimunitními chorobami, ale nyní se předpokládá jejich vlastní efektorové a regulační funkce. Na základě nedávných důkazů je třeba poznamenat, že mezi pomocnými T lymfocyty existuje funkční plasticita. Studie u myší prokázala schopnost Th17 lymfocytu transformovat do Th1 lymfocytu in vivo.[8] Následná studie prokázala rozsáhlou plasticitu pomocných T lymfocytů pro člověka.[9]
Paměťové T lymfocyty
Historicky, paměťové T lymfocyty patřily buď do subtypu efektorové paměti nebo subtypu centrální paměti, každý z nich měl odlišnou sadu povrchových buněčných markerů.[10] T lymfocyty centrální paměti sídlí v lymfatických uzlinách, zatímco T lymfocyty efektorové paměti postrádají C-C chemokinový receptor typu 7 (CCR7) a L-selektin (CD62L), což jim brání v přesunu do lymfatických uzlin.
Dnes jsou známy další populace paměťových T lymfocytů, ty zahrnují v tkáních sídlící paměťové T lymfocyty (Trm) a virtuální paměťové T lymfocyty[11]. Sjednocujícím znakem všech paměťových T lymfocytů je jejich dlouhodobé přežívání a schopnost rychle narůstat do velkého množství efektorových T lymfocytů, v případě opětovného setkání s dříve rozpoznaným antigenem. Tímto způsobem poskytují imunitnímu systému "paměť" proti dříve rozpoznaným patogenům.
Role v onemocnění
Vzhledem k různorodým rolím a důležitosti pomocných T lymfocytů v imunitním systému není překvapením, že tyto buňky ovlivňují imunitní odpověď proti chorobám. Zdá se, že příležitostně dělají chyby nebo vytvářejí odpovědi, které by se daly označit jako neprospěšné. V nejhorším možném případě, velmi vzácně, mohou pomocné T lymfocyty způsobit poškození až smrt hostitele.
Přecitlivělost
Úlohou imunitního systému je dosáhnout rovnováhy v citlivosti, aby byl schopen reagovat na cizí antigeny, aniž by reagoval na antigeny tělu vlastní. Když imunitní systém reaguje na velmi nízké hladiny antigenu, na které obvykle neodpovídá, může dojít ke vzniku přecitlivělosti. Tato přecitlivělost je považována za příčinu alergií a některých autoimunitních chorob.
Reakce na přecitlivělost můžeme rozdělit do čtyř typů (někdy označované jako Imunopatologické reakce):
- Typ 1 zahrnuje běžné imunitní poruchy jako astma, alergická rýma (senná rýma), ekzém, urtikárie (kopřivka) a anafylaktický šok. Všechny tyto reakce jsou spojeny s protilátkovou odpovědí typu IgE, které vyžadují Th2 imunitní odpověď. Preventivní léčba (kortikosteroidy) se zaměřuje na potlačení žírných buněk a dalších buněk spojených s alergií. T lymfocyty nehrají primární úlohu během vlastní zánětlivé odpovědi. Důležité je poznamenat, že označení jednotlivých typů neodpovídá typům pomocných T lymfocytů.
- Typ 2 a Typ 3 zahrnují komplikace spojené s autoimunitními nebo nízko afinními protilátkami. V obou typech reakcí se T lymfocyty podílí na tvorbě auto-specifických protilátek, ačkoli některé reakce Typu 2 by byly považovány za fyziologickém ve zdravém imunitním systému (například reakce proti Rh faktoru během těhotenství je normální imunitní odpovědí proti antigenům plodu). Porozumění role pomocných T lymfocytů v případě těchto reakcí je omezené, ale obecně se předpokládá, že Th2 cytokiny podporují tyto poruchy, například u choroby Systémový lupus erythematodes (SLE).
- Typ 4, také známý jako přecitlivělost opožděného typu, je způsobený nadměrnou stimulací imunitních buněk, běžně lymfocytů a makrofágů, která vede ke chronické zánětlivé odpovědi a uvolnění cytokinů. V tomto alergickém typu nehrají protilátky roli. T lymfocyty hrají důležitou roli v Typu 4, protože dochází k jejich aktivaci vůči stimulu a také podporují aktivaci dalších buněk, především makrofágů, skrze Th1 cytokiny
Další buněčné přecitlivělosti zahrnují autoimunitní choroby zprostředkované cytotoxickými T lymfocyty, podobným jevem je odmítnutí transplantánu. Pomocné T lymfocyty jsou součástí vývoje těchto chorob, aby byl umožněn vznik dostatečného množství cytotoxických T lymfocytů, je potřeba IL-2, který je dodáván CD4+ T lymfocyty. CD4+ buňky mohou prostřednictvím IFN-γ stimulovat i další buňky, jako jsou NK buňky a makrofágy, a povzbuzovat tyto cytotoxické buňky k usmrcení hostitelských buněk.
Mechanismus účinku cytotoxických T lymfocytů během autoimunit je téměř totožný s jejich reakcí proti virům, některé viry jsou proto spojeny se vznikem některých autoimunitních chorob jako je Diabetes mellitus prvního typu. Buněčné autoimunity se vyskytují v důsledku selhání rozpoznávání vlastních (hostitelských) antigenů, imunitní systém pak rozeznává vlastní antigeny jako cizí. V důsledku toho CD8+ lymfocyty považují hostitelskou buňku, prezentující autoantigen, za infikovanou a všechny hostitelské buňky ničí (v případě odmítnutí transplantátu, se toto týká buněk transplantátu prezentujících tělu cizí antigen).
Mnoho autoimunitních chorob je komplexních, dobře známý příklad je revmatoidní artritida, kde se patologie účastní jak protilátky, tak imunitní buňky. Obecně není imunologie většiny autoimunitních chorob dobře pochopena.
HIV infekce
Nejlepší příklad důležitosti CD4+ T lymfocytů je demonstrován infekcí viru lidské imunodeficience (HIV). Hlavním cílem HIV infekce jsou CD4+ T lymfocyty, ale může infikovat i jiné buňky exprimující molekulu CD4, například makrofágy a dendritické buňky (oba typy buněk exprimují CD4 v nízkých hladinách).
Bylo navrženo, že v průběhu nesymptomatické fáze HIV infekce, má virus poměrně nízkou afinitu k T lymfocytům (a má vyšší afinitu k makrofágům), což má za následek pomalou míru zabíjení CD4+ T lymfocytů imunitním systémem. To je zpočátku kompenzováno produkcí nových pomocných T lymfocytů z thymu (původem z kostní dřeně). Jakmile se však virus stane lymfotropním, začne mnohem efektivněji napadat CD4+ T lymfocyty (pravděpodobně díky změně koreceptorů, na které se během infekce váže) a imunitní systém je ohrožen. Je třeba poznamenat, že nedávné studie naznačují, že pouze ~ 5% CD4+ T lymfocytů infikovaných virem HIV jsou permisivní a stanou se produktivně infikovanými. Více než 95% CD4+ T lymfocytů, které zemřou, podstoupí abortivní infekci HIV[12]. Buněčná smrt je spuštěna, když hostitelská buňka rozpozná přítomnost HIV viru, což vede k aktivaci kaspázy 1 a následně pyroptóze (vysoce zánětlivá forma programované buněčné smrti).[13][14]
V tomto okamžiku nastává chronický zánět a funkční hladiny CD4+ T lymfocytů se začínají snižovat, časem až do bodu, kdy je populace CD4+ buněk příliš malá, aby byla schopna rozpoznat celou řadu antigenů, které by mohly být potenciálně detekovány. Vyčerpání CD4+ T lymfocytů a vývoj chronického zánětu jsou typické procesy v patogenezi HIV, které vedou k vývoji získaného syndromu imunitní nedostatečnosti (AIDS). Snížení počtu CD4+ T lymfocytů v krvi na méně než 200 buněk/μl, umožňuje různým patogenům uniknout rozpoznání T lymfocyty a vede k oportunním infekcím.
Díky své závislosti na CD4+ T lymfocytech jsou v důsledku AIDS postiženy především dvě složky imunitního systému:
- CD8+ T lymfocyty nejsou efektivně stimulovány během AIDS fáze HIV infekce, což činí pacienty velmi náchylné k většině virů, včetně samotného HIV. Tento pokles v zabíjení infikovaných CD4+ T lymfocytů vede k tomu, že virus je produkován delší dobu (infikované CD4+ T buňky nejsou dostatečně rychle zničeny), zvyšuje se proliferace viru a urychluje vývoj onemocnění.
- Důsledkem selhání funkce pomocných T lymfocytů dochází k výraznému snížení procesu izotypového přesmyku protilátek. Imunitní systém ztrácí schopnost zlepšit afinitu svých protilátek a nedokáže generovat B lymfocyty produkující protilátky tříd IgG a IgA. Tyto účinky jsou primárně způsobeny ztrátou pomocných T lymfocytů schopných správně interagovat s B lymfocyty. Dalším příznakem AIDS je snížení hladin protilátek kvůli nedostatku Th2 cytokinů (a nedostatku interakcí pomocných T lymfocytů). Tyto komplikace vedou ke zvýšené náchylnosti k bakteriálním infekcím, především v oblastech těla, které nejsou přístupné IgM protilátkám.
Jestliže pacient nereaguje na (nebo neobdrží) léčbu proti viru HIV, podléhá obvykle rakovinám nebo infekcím, imunitní systém nakonec dosáhne bodu, kdy již není schopen čelit infekcím
Reference
V tomto článku byl použit překlad textu z článku T helper cell na anglické Wikipedii.
- VAN DEN BROEK, Theo; BORGHANS, José A. M.; VAN WIJK, Femke. The full spectrum of human naive T cells. Nature Reviews. Immunology. 2018-6, roč. 18, čís. 6, s. 363–373. PMID: 29520044. Dostupné online [cit. 2019-02-13]. ISSN 1474-1741. DOI 10.1038/s41577-018-0001-y. PMID 29520044.
- VAN DEN BROEK, Theo; DELEMARRE, Eveline M.; JANSSEN, Willemijn J.M. Neonatal thymectomy reveals differentiation and plasticity within human naive T cells. The Journal of Clinical Investigation. Roč. 126, čís. 3, s. 1126–1136. PMID: 26901814 PMCID: PMC4767338. Dostupné online [cit. 2019-02-13]. ISSN 0021-9738. DOI 10.1172/JCI84997. PMID 26901814.
- ZHU, Jinfang; PAUL, William E. CD4 T cells: fates, functions, and faults. Blood. 2008-09-01, roč. 112, čís. 5, s. 1557–1569. PMID: 18725574 PMCID: PMC2518872. Dostupné online [cit. 2019-02-21]. ISSN 0006-4971. DOI 10.1182/blood-2008-05-078154. PMID 18725574.
- ScienceDirect. www.sciencedirect.com [online]. [cit. 2019-02-21]. Dostupné online.
- MAVERAKIS, Emanual; KIM, Kyoungmi; SHIMODA, Michiko. Glycans In The Immune system and The Altered Glycan Theory of Autoimmunity: A Critical Review. Journal of autoimmunity. 2015-2, roč. 0, s. 1–13. PMID: 25578468 PMCID: PMC4340844. Dostupné online [cit. 2019-02-21]. ISSN 0896-8411. DOI 10.1016/j.jaut.2014.12.002. PMID 25578468.
- RABINOVICH, Gabriel A.; BAUM, Linda G.; RILEY, Eleanor M. Differential glycosylation of TH1, TH2 and TH-17 effector cells selectively regulates susceptibility to cell death. Nature Immunology. 2007-08, roč. 8, čís. 8, s. 825–834. Dostupné online [cit. 2019-02-21]. ISSN 1529-2916. DOI 10.1038/ni1482. (anglicky)
- WEAVER, Casey T.; MURPHY, Kenneth M.; MURPHY, Theresa L. Interleukin 17–producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nature Immunology. 2005-11, roč. 6, čís. 11, s. 1123–1132. Dostupné online [cit. 2019-02-21]. ISSN 1529-2916. DOI 10.1038/ni1254. (anglicky)
- HIROTA, K.; DUARTE, J.H.; VELDHOEN, M. Fate mapping of interleukin 17-producing T cells in inflammatory responses. Nature immunology. 2011-3, roč. 12, čís. 3, s. 255–263. PMID: 21278737 PMCID: PMC3040235. Dostupné online [cit. 2019-02-21]. ISSN 1529-2908. DOI 10.1038/ni.1993. PMID 21278737.
- LARSEN, Martin; ARNAUD, Laurent; HIÉ, Miguel. Multiparameter grouping delineates heterogeneous populations of human IL-17 and/or IL-22 T-cell producers that share antigen specificities with other T-cell subsets. European Journal of Immunology. 2011-9, roč. 41, čís. 9, s. 2596–2605. PMID: 21688259. Dostupné online [cit. 2019-02-21]. ISSN 1521-4141. DOI 10.1002/eji.201041131. PMID 21688259.
- SALLUSTO, F.; LENIG, D.; FÖRSTER, R. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature. 1999-10-14, roč. 401, čís. 6754, s. 708–712. PMID: 10537110. Dostupné online [cit. 2019-02-21]. ISSN 0028-0836. DOI 10.1038/44385. PMID 10537110.
- MARUSINA, Alina I.; ONO, Yoko; MERLEEV, Alexander A. CD4+ virtual memory: Antigen-inexperienced T cells reside in the naïve, regulatory, and memory T cell compartments at similar frequencies, implications for autoimmunity. Journal of Autoimmunity. 02 2017, roč. 77, s. 76–88. PMID: 27894837 PMCID: PMC6066671. Dostupné online [cit. 2019-02-21]. ISSN 1095-9157. DOI 10.1016/j.jaut.2016.11.001. PMID 27894837.
- DOITSH, Gilad; CAVROIS, Marielle; LASSEN, Kara G. Abortive HIV infection mediates CD4 T cell depletion and inflammation in human lymphoid tissue. Cell. 2010-11-24, roč. 143, čís. 5, s. 789–801. PMID: 21111238 PMCID: PMC3026834. Dostupné online [cit. 2019-02-21]. ISSN 1097-4172. DOI 10.1016/j.cell.2010.11.001. PMID 21111238.
- DOITSH, Gilad; GALLOWAY, Nicole L. K.; GENG, Xin. Cell death by pyroptosis drives CD4 T-cell depletion in HIV-1 infection. Nature. 2014-01-23, roč. 505, čís. 7484, s. 509–514. PMID: 24356306 PMCID: PMC4047036. Dostupné online [cit. 2019-02-21]. ISSN 1476-4687. DOI 10.1038/nature12940. PMID 24356306.
- MONROE, Kathryn M.; YANG, Zhiyuan; JOHNSON, Jeffrey R. IFI16 DNA sensor is required for death of lymphoid CD4 T cells abortively infected with HIV. Science (New York, N.Y.). 2014-01-24, roč. 343, čís. 6169, s. 428–432. PMID: 24356113 PMCID: PMC3976200. Dostupné online [cit. 2019-02-21]. ISSN 1095-9203. DOI 10.1126/science.1243640. PMID 24356113.
Externí odkazy
 Obrázky, zvuky či videa k tématu Pomocný T-lymfocyt na Wikimedia Commons
Obrázky, zvuky či videa k tématu Pomocný T-lymfocyt na Wikimedia Commons