TLR3
Toll-like receptor 3 (TLR3) je protein, který je u lidí kódován genem TLR3.[1] TLR3 je transmembránový receptor patřící do rodiny toll-like receptorů, které patří mezi pattern-recognition receptory (PRR). Podílí se na imunitní obraně rozpoznáváním konzervovaných struktur typických pro patogenní mikroorganismy, tvz. PAMPs (pathogen-associated molecular patterns) s následnou signalizací a aktivací vrozeného imunitního systému. Jeho nejznámějším ligandem je dvouvláknová RNA, charakteristická pro různé virové patogeny. Mezi jeho další ligandy patří syntetické sloučeniny známé jako poly I-C (polyinosin:polycytidylované kyseliny), používané k simulaci virových infekcí.
TLR3 byl zároveň označen jako CD283 (diferenciační skupina 283).
Funkce
TLR3 kódovaný tímto genem, patří do rodiny toll-like receptorů (TLR), která hraje zásadní roli při rozpoznávání patogenů a aktivaci přirozené imunity. TLR jsou odvozené od proteinu toll poprvé popsaného u octomilky Drosophily, kde kromě kontroly dorzoventrální osy embrya, rovněž reaguje na cizorodé molekuly a vzhledem k vysoké konzerovanosti sdílí se savčími TLR jisté strukturální a funkční podobnosti. Rozeznávají molekulární vzory asociované s patogeny (PAMP), které jsou exprimovány na infekčních agens, a zprostředkovávají produkci cytokinů nezbytných pro vývoj účinné imunity. Různé TLR vykazují rozdílné vzory exprese. Receptor TLR3 je nejhojněji exprimován v placentě, slinivce, plicích, srdci, játrech, mozku a v dendritické subpopulaci leukocytů.[2] Rozeznává dsRNA asociovanou s virovou infekcí a indukuje aktivaci signalizační dráhy IRF3 a NF-κB.[3] Na rozdíl od jiných TLR používá TLR3 jako jediný adaptér TRIF. IRF3 nakonec indukuje produkci interferonů typu I, které hrají důležitou úlohu při obraně hostitele proti virům.[4]
TLR3 rozeznávají dsRNA, formu genetické informace přítomné některých virů, jako jsou reoviry. Kromě toho existuje ještě efemérní forma siRNA, účastnící se RNA interference. TLR 3 po rozeznání ligandu indukuje aktivaci IRF3 s následným zvýšením produkce interferonů typu I, které signalizují jiným buňkám, aby zvýšily svou antivirovou obranu. Dvouřetězcová RNA je také rozpoznána cytoplazmatickými receptory RIG-I a MDA-5.[5]
Bylo prokázáno, že TLR3 hraje obranou úlohu v myších modelech u onemocnění aterosklerózy,[6] mozkové ischemii a reaktivní astrogliózy.[7][8] Dále bylo zjištěno, že aktivace TLR3 podporuje regeneraci vlasových folikulů při hojení kožních ran.[9] Kromě toho aktivátory TLR3 vykazují účinky na lidské vaskulární buňky.[6]
Struktura
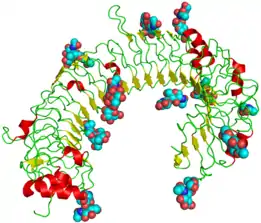
Struktura TLR3 byla objevena v roce 2005 nezávisle dvěma výzkumnými skupinami, v červnu vědci ze The Scripps Research Institute a v srpnu americkou skupinou v čele s J.Bellovou.[10][11] TLR3 je integrální membránový receptor, skládající se z extracelulární, transmembránové a intracelulární domény.[12]
- Extracelulární doména je solenoid ve tvaru podkovy složený z 23 repetic bohatých na leucin tzv. LRR (leucine rich repeats), kde LRR tvoří jednu otáčku solenoidu. Extracelulární doména je na N- a C- terminálním konci zakončená Cap doménou bohatou na cystein. Konkávní, tedy plocha proteinu vyklenutá dovnitř, je bohatá na potenciální N-glykosylační místa a je pravděpodobně silně glykosylována[10], zatímco konvexní strana (vně podkovy) je bez glykosylace. Dle předpokladu má extracelulární doména 15 vazebných míst.
- Transmembránová doména jednou šroubovicí procházející skrz membránu.
- Intracelulární (cytoplazmatická) doména obsahuje oblast CYT a a doménu TIR, která se účastní vazby na TRIF.
Velká část povrchu receptoru TLR3 je pokryta molekulami sacharidů, což z něj dělá glykoprotein, přesto může snadno krystalizovat, jak bylo potvrzeno strukturní analýzou rentgenové krystalografie. V této krystalové struktuře může dimerizovat. C-terminální konce extracelulární domény se dotýkají sousední podkovy a tvoří tak "dimer" dvou podkov, proto hraje důležitou roli v dimerizaci a signalizaci.[13]
Komplex TLR3-dsRNA
Výzkumná skupina v čele s Chloe, kteří publikovali strukturu v červnu 2005 předpokládali, že dsRNA by se mohla vázat na konvexním povrchu TLR3, protože tato oblast je bez glykanů, což umožňuje pro záporně nabitou dsRNA vázat se na kladně nabité aminokyselinové zbytky extracelulární domény TLR.[10]
Bellová a kol. navrhli, že dsRNA by se mohla naopak vázat na konkávním povrchu. Předpoklad byl postaven na skutečnosti, že během krystalizace molekuly síranu z krystalizačního média vážou zbytky v LRR 11 a LRR 20 extracelulární domény TLR3. A sírany sdílí stejné atomové uspořádání jako fosfátové skupiny přítomné v dsRNA, tedy by se mohly být schopny vázat jedno nebo obě sulfátová vazebná místa.[11]
Extracelulární doména TLR3 v roztoku existuje jako monomer a k dimerizaci dochází po navázání ligandu. dsRNA interaguje s N- a C-terminálními místy na laterální straně konvexního povrchu extracelulární domény TLR3 a to konkrétně s LRR20 - lokalizováno na bez glykanové laterální straně. TLR3 po navázání ligandu následně vytvoří dimer ze dvou podkov solenoidů.[14]
Signalizace
TLR3 na rozdíl od ostatních členů TLR rodiny nepoužívá myeloidní diferenciační faktor 88 (MyD88) jako signalizační adaptér. Po rozeznání ligandu a dimerizaci dochází k náboru adaptorových molekul, které jsou nezbytné pro přenos signálu uvnitř buňky. TLR3 zprostředkovává signály prostřednictvím adaptorového proteinu TICAM-1/TRIF.[15] Signalizační dráha indukovaná přes TICAM-1 aktivuje transkripční faktory jako nukleárního faktoru kappa B (NF-κB), IRF-3 a AP-1 což vede k produkci cytokinů INF typu I.
TIR doména intracelulární části TLR3 je zodpovědná za nábor adaptorových molekul a následnou signalizaci a to prostřednictvím alaninového zbytku Ala795H v B-B smyčce, který je nahrazen histidinovým zbytkem.
Exprese
TLR3 je exprimován převážně intracelulárně, kde má vhodné podmínky k setkání s ligandy uvolněnými z pohlcených mikroorganismů. TLR3 je hojně exprimován myeloidními dendritickými buňkami, střevním epitelem, polymorfonukleárními leukocyty, B lymfocytech a T lymfocytech, NK buňkami, ale ne v monocytech.[16][17] V makrofázích, DC, fibroblastech a epiteliálních buňkách je exprese TLR3 up-regulována virovou infekcí a exogenním přidáním poly (I:C) nebo cytokinem IFN typu I.[18] Kromě toho i v astrocytech se může po stimulaci poly (I:C) zvýšit exprese TL3.[19]
Ligandy
- Antagonisté
Bylo zjištěno, že TLR3 interaguje s:
- CU-CPT4a
TLR3 rozpoznává dsRNA a poly (I:C) a jelikož je exprimován na intracelulárních membránách, ligandy jsou často transfekovány do buněk pomocí kationtových lipozomů jako je lipofectin.
Reference
V tomto článku byl použit překlad textu z článku TLR3 na anglické Wikipedii.
- ROCK, F. L.; HARDIMAN, G.; TIMANS, J. C. A family of human receptors structurally related to Drosophila Toll. Proceedings of the National Academy of Sciences. 1998-01-20, roč. 95, čís. 2, s. 588–593. Dostupné online [cit. 2022-02-06]. ISSN 0027-8424. DOI 10.1073/pnas.95.2.588.
- MATSUMOTO, Misako. Innate Immune Recognition of Microbial Components by Toll-Like Receptors.. Seibutsu Butsuri. 2002, roč. 42, čís. 6, s. 295–300. Dostupné online [cit. 2022-02-06]. ISSN 0582-4052. DOI 10.2142/biophys.42.295. (anglicky)
- KAWAI, Taro; AKIRA, Shizuo. TLR signaling. Seminars in Immunology. 2007-02, roč. 19, čís. 1, s. 24–32. Dostupné online [cit. 2022-02-06]. ISSN 1044-5323. DOI 10.1016/j.smim.2006.12.004.
- Toll-Like Receptor Family Gene. [s.l.]: Qeios Dostupné online.
- ALEXOPOULOU, Lena; HOLT, Agnieszka Czopik; MEDZHITOV, Ruslan. Recognition of double-stranded RNA and activation of NF-κB by Toll-like receptor 3. Nature. 2001-10, roč. 413, čís. 6857, s. 732–738. Dostupné online [cit. 2022-02-06]. ISSN 0028-0836. DOI 10.1038/35099560.
- COLE, J.E.; NAVIN, T.; CROSS, A.J. 11 THE EXPRESSION OF TLR3 IS INCREASED IN ATHEROSCLEROSIS AND MEDIATES PROTECTION AGAINST LESION DEVELOPMENT AND WEIGHT GAIN IN APOE−/− MICE. Atherosclerosis Supplements. 2011-06, roč. 12, čís. 1, s. 3. Dostupné online [cit. 2022-02-06]. ISSN 1567-5688. DOI 10.1016/s1567-5688(11)70012-5.
- GUO, Wenxi; LI, Xiaoyi; CHEN, Mengxiao. Electrochemical Cathodic Protection Powered by Triboelectric Nanogenerator. Advanced Functional Materials. 2014-08-26, roč. 24, čís. 42, s. 6691–6699. Dostupné online [cit. 2022-02-06]. ISSN 1616-301X. DOI 10.1002/adfm.201401168.
- LI, Yang; XU, Xu-Lin; ZHAO, Dan. TLR3 ligand Poly IC Attenuates Reactive Astrogliosis and Improves Recovery of Rats after Focal Cerebral Ischemia. CNS Neuroscience & Therapeutics. 2015-10-23, roč. 21, čís. 11, s. 905–913. Dostupné online [cit. 2022-02-06]. ISSN 1755-5930. DOI 10.1111/cns.12469.
- NELSON, Amanda M.; REDDY, Sashank K.; RATLIFF, Tabetha S. dsRNA Released by Tissue Damage Activates TLR3 to Drive Skin Regeneration. Cell Stem Cell. 2015-08, roč. 17, čís. 2, s. 139–151. Dostupné online [cit. 2022-02-06]. ISSN 1934-5909. DOI 10.1016/j.stem.2015.07.008.
- CHOE, Jungwoo; KELKER, Matthew S.; WILSON, Ian A. Crystal Structure of Human Toll-Like Receptor 3 (TLR3) Ectodomain. Science. 2005-07-22, roč. 309, čís. 5734, s. 581–585. Dostupné online [cit. 2022-02-06]. ISSN 0036-8075. DOI 10.1126/science.1115253.
- BELL, J. K.; BOTOS, I.; HALL, P. R. The molecular structure of the Toll-like receptor 3 ligand-binding domain. Proceedings of the National Academy of Sciences. 2005-08-02, roč. 102, čís. 31, s. 10976–10980. Dostupné online [cit. 2022-02-06]. ISSN 0027-8424. DOI 10.1073/pnas.0505077102. PMID 16043704. (anglicky)
- BELL, Jessica K.; MULLEN, Gregory E. D.; LEIFER, Cynthia A. Leucine-rich repeats and pathogen recognition in Toll-like receptors. Trends in Immunology. 2003-10, roč. 24, čís. 10, s. 528–533. PMID 14552836. Dostupné online [cit. 2022-02-06]. ISSN 1471-4906. DOI 10.1016/s1471-4906(03)00242-4. PMID 14552836.
- TAKADA, Eiko; OKAHIRA, Satoyo; SASAI, Miwa. C-terminal LRRs of human Toll-like receptor 3 control receptor dimerization and signal transmission. Molecular Immunology. 2007-07, roč. 44, čís. 15, s. 3633–3640. Dostupné online [cit. 2022-02-06]. DOI 10.1016/j.molimm.2007.04.021. (anglicky)
- LIU, Lin; BOTOS, Istvan; WANG, Yan. Structural Basis of Toll-Like Receptor 3 Signaling with Double-Stranded RNA. Science. 2008-04-18, roč. 320, čís. 5874, s. 379–381. Dostupné online [cit. 2022-02-06]. ISSN 0036-8075. DOI 10.1126/science.1155406. PMID 18420935. (anglicky)
- OSHIUMI, Hiroyuki; MATSUMOTO, Misako; FUNAMI, Kenji. TICAM-1, an adaptor molecule that participates in Toll-like receptor 3–mediated interferon-β induction. Nature Immunology. 2003-02, roč. 4, čís. 2, s. 161–167. Dostupné online [cit. 2022-02-06]. ISSN 1529-2908. DOI 10.1038/ni886. (anglicky)
- CARIO, Elke; PODOLSKY, Daniel K. Differential Alteration in Intestinal Epithelial Cell Expression of Toll-Like Receptor 3 (TLR3) and TLR4 in Inflammatory Bowel Disease. Infection and Immunity. 2000-12, roč. 68, čís. 12, s. 7010–7017. Dostupné online [cit. 2022-02-06]. ISSN 0019-9567. DOI 10.1128/IAI.68.12.7010-7017.2000. PMID 11083826. (anglicky)
- MUZIO, Marta; BOSISIO, Daniela; POLENTARUTTI, Nadia. Differential Expression and Regulation of Toll-Like Receptors (TLR) in Human Leukocytes: Selective Expression of TLR3 in Dendritic Cells. The Journal of Immunology. 2000-06-01, roč. 164, čís. 11, s. 5998–6004. PMID 10820283. Dostupné online [cit. 2022-02-06]. ISSN 0022-1767. DOI 10.4049/jimmunol.164.11.5998. PMID 10820283. (anglicky)
- VERCAMMEN, Elisabeth; STAAL, Jens; BEYAERT, Rudi. Sensing of Viral Infection and Activation of Innate Immunity by Toll-Like Receptor 3. Clinical Microbiology Reviews. 2008-January. Dostupné online [cit. 2022-02-06]. DOI 10.1128/CMR.00022-07. PMID 18202435. (EN)
- CARTY, Michael; BOWIE, Andrew. Toll-like receptor 3. New Drugs and Targets for Asthma and COPD. 2010, roč. 39, s. 73–79. Dostupné online [cit. 2022-02-06]. DOI 10.1159/000320802. (english)