RecQ helikáza
RecQ helikáza je označení pro vysoce konzervovanou rodinu helikáz, které jsou díky své nepostradatelné roli v mnoha procesech souvisejících s DNA považovány za tzv. „ochránce“ genomu, protože se podílí na udržování stability genetické informace a zároveň potlačují vznik nádorového bujení.[1][2] [3]
Zatímco u bakterií a kvasinek byla identifikována pouze jedna RecQ helikáza, v lidských buňkách bylo objeveno dokonce pět RecQ homologních proteinů – RECQ1[4][5], BLM (RECQ2) [6], WRN (RECQ3)[7], RECQ4 (RTS) a RECQ5 [8]. Nepostradatelná biologická role RecQ helikáz je navíc podtržena tím, že mutace v genech BLM, WRN a RECQ4 jsou spojeny s velmi zákeřnými dědičnými onemocněními: Bloomovým (BS), Wernerovým (WS) a Rothmund-Thomsonovým syndromem (RTS). Tato genetická onemocnění jsou spojena s vyšší náchylností k rakovině, předčasným stárnutím a chromozomovou nestabilitou.[9][10][11]
Struktura a biochemická aktivita
Podle experimentálních a bioinformatických analýz lze uvnitř polypeptidového řetězce RecQ helikáz rozlišit několik konzervovaných domén, určujících zároveň aktivitu jednotlivých proteinů. Hlavní domény RecQ helikáz jsou tři: helikázová, RecQ karboxy-terminální (RQC) a RNázováD C-terminální (HRDC) doména [12].
- Helikázová doména s rozsahem 450 aminokyselin zahrnuje motiv pro vazbu ATP, který je hlavním energetickým zdrojem pro samotnou helikázovou aktivitu. BLM a WRN, které patří mezi nejvíce prostudované RecQ proteiny, umí rozvinout nejrůznější DNA struktury zahrnující 3‘-převis, bublinové struktury, Y-formy, D-smyčky, křížovou DNA strukturu zvanou Holliday junctions (HJ) nebo G-kvadruplexové struktury [13][14][15].
- RQC doména je důležitá pro strukturní celistvost proteinu (viz strukturní alignment) a také pro vazbu dvouřetězcové DNA [16][17].
Tato doména bude mít také pravděpodobně roli při zprostředkování proteinových interakcí RecQ helikáz s jinými proteiny.[18]
- Poslední HRDC doména je nejméně konzervovaná, nicméně pomocí rentgenové krystalografie bylo zjištěno, že je důležitá pro vazbu jednořetězcové DNA. Domény RQC a HRDC, důležité pro vazbu k DNA, u některých členů RecQ helikázové rodiny chybějí, např. RECQ4 postrádá obě domény [19][20]
Kromě výše popsaných domén se mohou RecQ helikázy mezi sebou lišit dalšími doménami, které mohou sdílet homologii s jinými proteiny a tím se liší i jejich vlastnosti a funkce v buňce. Příkladem může být WRN helikáza s exodoménou na N-konci, která zajišťuje exonukleázovou aktivitu WRN proteinu. [21] Lidské RECQ helikázy se vyznačují nejen klasickou helikázovou aktivitou, tedy schopností oddělovat od sebe dva řetězce DNA za přítomnosti ATP. Na druhou stranu vykazují také velmi silnou tzv. annealing aktivitu, tj. zprostředkovávat nasedání komplementárních jednořetězcových úseků DNA do výsledného duplexu. [22]
Funkce
Buněčný cyklus zahrnuje několik kroků, z nichž každý musí být efektivně a časově dokončen než započne krok následující. Během S-fáze buněčného cyklu buňka musí precizně duplikovat své chromozomy, které pak následně musí velmi přesně rozdělit během M-fáze. Avšak během S-fáze se může vyskytnout několik překážek, které mohou nejen ovlivnit buněčný cyklus, ale také mohou vést v následující M-fázi k chromozomové nestabilitě, která bývá obvykle příčinou vzniku některých neurologických onemocnění nebo nádorového bujení. [23]
Replikace DNA
Během replikace DNA se mohou vyskytnout překážky, které replikační mašinerie není schopná vyřešit, a proto dochází k předčasnému zastavení replikační vidlice [23] [24] . Problémy replikační vidlice mohou být způsobeny těmito překážkami:
- Adukty DNA, tedy pozměněné nukleotidy (nukleosidy nebo báze). Příčinou vzniku aduktů jsou obvykle elektrofilní organické látky, které vstupují do organismu z vnějšího prostředí anebo vznikají jako reaktivní metabolické intermediáty, se schopností atakovat nukleofilní centra v nukleových kyselinách a tímto poškozovat genetickou informaci.
- Jednořetězcové nebo dvouřetězcové zlomy, které vznikají především po expozici ionizujícím zářením, UV zářením (viz ultrafialové záření), genotoxickými chemikáliemi, ale také produkty vnitřního oxidativního metabolismu buňky.
- Kovalentně vázaný protein na DNA, který replikační helikáza není schopná odstranit. Problémy může také způsobovat setkání procesů replikace a transkripce.
- Různé intramolekulární sekundární struktury ssDNA (např. G-quadruplex) ve vedoucím řetězci rovněž vedou k zastavení replikační vidlice.
- Topologický stres, kdy se potkají dvě proti sobě jdoucí replikační vidlice. Tento problém je často nevyhnutelný rys eukaryotické replikace, ve které je obousměrná replikace iniciována z více počátků replikace podél každého chromozomu.
RecQ helikázy se na odstranění replikačních překážek podílí mnoha způsoby. Jedna z nejdůležitějších funkcí je restart zastavené replikační vidlice. Výskyt poškození na vedoucím řetězci (DNA adukt) vede k zastavení replikační vidlice, avšak BLM a WRN podporují tzv. regresi replikační vidlice, což znamená oddělení nově nasyntetizovaných DNA vláken a jejich spojení, čímž vzniká čtyřcestná Holliday struktura [25] [26] . Vzniklá Hollidayova struktura pak může migrovat zpět za pomocí zejména RECQ1, BLM nebo WRN, a tím dochází k obnovení replikační vidlice. [27] [28] [29] Alternativně může být Hollidayova struktura rozštěpena strukturně specifickými endonukleázami, což vede ke kolapsu replikační vidlice, která pak musí být opravena pomocí procesu homologní rekombinace (viz níže. [30] [31] [32] RecQ helikázy (zejména BLM a WRN) jsou schopné také díky své enzymatické aktivitě rozvíjet různé DNA struktury (G-quadruplexy, různé smyčky a vlásenky), které vznikají jako replikační „překážky“ vedoucího řetězce. [33] Navíc BLM také zastává důležitou úlohu při řešení topologického stresu, který vzniká při setkání dvou protichůdných replikačních vidlic. Přesná molekulární podstata je jen velmi málo objasněna, avšak důležitou roli při řešení tohoto problému hraje komplex zahrnující BLM-TOPOIIIα(DNA Topoizomeráza IIIα)-RMI1-RMI2, proteiny, které jsou schopny topologický stres uvolnit. [34]
DNA oprava
Jak bylo popsáno výše, DNA je neustále poškozována exogenním i endogenním působením, které způsobují mnoho různých typů poškození v DNA. V důsledku těchto rozdílných typů poškození (depurinace, deaminace, thyminové dimery, zlomy DNA aj.) se v průběhu evoluce vyvinula celá řada opravných mechanismů zodpovědná za jejich odstranění. RecQ helikázy i zde zastávají mnoho úloh napříč téměř všemi mechanismy. Hlavní DNA opravné mechanismy tvoří:
| Mechanismus opravy | Vznik poškození |
|---|---|
| Nukleotidová excisní oprava (NER) | Poškození, která narušují helikální dvoušroubovici DNA,
jako jsou rozměrné bázové adukty a fotoprodukty UV záření (př. thyminové dimery). |
| Homologní rekombinace (HR) | dvouřetězcové zlomy DNA, zastavené replikační vidlice |
| Nehomologní spojování konců (NHEJ) | dvouřetězcové zlomy DNA |
| Oprava nesprávného párování (MMR) | malé inzerce, delece a nesprávné zařazení bází během replikace a rekombinace. |
| Básová excisní oprava (BER) | Abnormální DNA báze, jednoduché bázové adukty, jednořetězcové zlomy DNA, při oxidativních poškozeních nebo při neúspěšné aktivitě topoizomerázy I,(např. báze chemicky poškozené alkylací, deaminací nebo oxidací). |
V rámci této práce[který?] se primárně zaměříme na mechanizmus opravy dvouřetězcových zlomů (DSB) pomocí homologní rekombinace. Oprava dvouřetězcových zlomů je esenciální pro život buňky, protože neschopnost opravy DSB může způsobovat aneuploidii a genomovou nestabilitu vedoucí k rakovině nebo buněčné smrti.
Homologní rekombinace
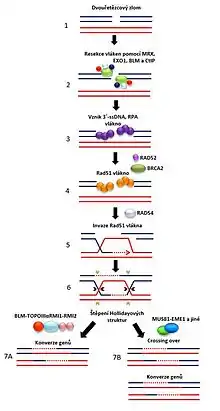
Homologní rekombinace (HR) je velmi precizní mechanismus zahrnující několik fází, kdy DSB je opraven na základě syntézy podle homologického úseku sesterské chromatidy nebo homologního chromozomu. V první fázi HR jsou konce DSB podrobeny resekci za vzniku 3‘ jednořetězcového úseku DNA (ssDNA). 3‘ ssDNA vlákno je ihned po resekci pokryto proteinem RPA (replikační protein A), který zabraňuje vzniku sekundárních struktur na ssDNA vlákně.[36] Následně dochází za pomocí rekombinačních mediatorů k výměně RPA za protein RAD51 a vzniku tzv. RAD51 presynaptického vlákna.[37] V druhé fázi HR je RAD51 presynaptické vlákno schopno vyhledat homologii za vzniku struktury zvané D-smyčka, ve které je pak donorová molekula DNA využita jako templát pro syntézu chybějící části.[38] Po DNA syntéze je nové vlákno zachyceno druhým koncem původního řetězce, což vede ke vzniku dvojité Hollidayovi struktury. HJ struktura musí být rozložena, aby byla dokončena rekombinace a dvě molekuly DNA se mohly oddělit. HJ struktury mohou být tedy štěpeny strukturně specifickými endonukleázami (např. MUS81-EME1) nebo rozloženy pomocí kombinované aktivity helikáz a topoizomeráz.[20][32]
RecQ helikázy zastávají důležité regulační funkce v průběhu homologní rekombinace. V první fázi HR se BLM helikáza přímo účastní resekce konců dvouřetězcového zlomu za vzniku 3’ ssDNA vlákna.[39] Několik studií prokázalo, že BLM, WRN a RECQ5 jsou schopny interagovat s proteinem RAD51 a vytlačovat jej z presynaptického vlákna, aby bylo zabráněno nevhodné nebo předčasné rekombinaci.[40][41] V poslední fázi HR se BLM-TOPOIIIα-RMI1-RMI2 komplex podílí na rozkladu dvojitých Hollidayových struktur.[20]
Onemocnění
S nesprávnou funkcí RecQ helikáz je spojeno několik genetických onemocnění.
| Onemocnění | Hlavní klinické znaky | Predispozice k rakovině |
|---|---|---|
| Bloomův syndrom | proporcionální malý vzrůst, imunodeficience, diabetes typ 2, mužská infertilita a ženská subfertilita, hyperpigmentace a hypopigmentace (kůže), k slunci citlivá obličejová erythema | většina typů, leukémie, rakoviny kůže, prsu a tračníku |
| Wernerův syndrom | různé znaky předčasného stárnutí: arterioskleróza, diabetes typ 2, osteoporóza, katarakt, šednutí a ztráta vlasů; retardace růstu v pubertě, kalcifikace měkké tkáně | převážně rakoviny mesenchymálního původu (měkká tkáň a jiné sarkomy), epitheliální sarkomy (melanomy) |
| Rothmund-Thomsonův syndrom | poikiloderma, skeletární abnormality, šedý zákal, řídnutí vlasů, dystrofické nehty a zuby | Nejčastěji osteosarkoma, často v dětství |
Reference
- [Nakayama H, Nakayama K, Nakayama R, Irino N, Nakayama Y, Hanawalt PC. Isolation and genetic characterization of a thymineless death-resistant mutant of Escherichia coli K12: identification of a new mutation (recQ1) that blocks the RecF recombination pathway. Mol Gen Genet 1984, 195: 474-480]
- [Mohaghegh P, Hickson ID. DNA helicase deficiencies associated with cancer predisposition and premature ageing disorders. Hum Mol Genet 2001, 10: 741-746]
- [Chu WK, Hickson ID. RecQ helicases: multifunctional genome caretakers. Nat Rev Cancer 2009, 9: 644-654]
- [Puranam KL, Blackshear PJ. Cloning and characterization of RECQL, a potential human homologue of the Escherichia coli DNA helicase RecQ. J Biol Chem 1994, 269: 29838-29845]
- [Seki M, Miyazawa H, Tada S, Yanagisawa J, Yamaoka T, Hoshino S, Ozawa K, et al. Molecular cloning of cDNA encoding human DNA helicase Q1 which has homology to Escherichia coli Rec Q helicase and localization of the gene at chromosome 12p12. Nucleic Acids Res 1994, 22: 4566-4573]
- [Ellis NA, Groden J, Ye TZ, Straughen J, Lennon DJ, Ciocci S, Proytcheva M, et al. The Bloom's syndrome gene product is homologous to RecQ helicases. Cell 1995, 83: 655-666]
- [Yu CE, Oshima J, Fu YH, Wijsman EM, Hisama F, Alisch R, Matthews S, et al. Positional cloning of the Werner's syndrome gene. Science 1996, 272: 258-262]
- [Kitao S, Ohsugi I, Ichikawa K, Goto M, Furuichi Y, Shimamoto A. Cloning of two new human helicase genes of the RecQ family: biological significance of multiple species in higher eukaryotes. Genomics 1998, 54: 443-452]
- [Ellis NA, German J. Molecular genetics of Bloom's syndrome. Hum Mol Genet 1996, 5 Spec No: 1457-1463]
- [Yu CE, Oshima J, Wijsman EM, Nakura J, Miki T, Piussan C, Matthews S, et al. Mutations in the consensus helicase domains of the Werner syndrome gene. Werner's Syndrome Collaborative Group. Am J Hum Genet 1997, 60: 330-341]
- [Kitao S, Shimamoto A, Goto M, Miller RW, Smithson WA, Lindor NM, Furuichi Y. Mutations in RECQL4 cause a subset of cases of Rothmund-Thomson syndrome. Nat Genet 1999, 22: 82-84]
- [Bernstein DA, Keck JL. Domain mapping of Escherichia coli RecQ defines the roles of conserved N- and C-terminal regions in the RecQ family. Nucleic Acids Res 2003, 31: 2778-2785]
- [Suzuki N, Shimamoto A, Imamura O, Kuromitsu J, Kitao S, Goto M, Furuichi Y. DNA helicase activity in Werner's syndrome gene product synthesized in a baculovirus system. Nucleic Acids Res 1997, 25: 2973-2978]
- [Karow JK, Chakraverty RK, Hickson ID. The Bloom's syndrome gene product is a 3'-5' DNA helicase. J Biol Chem 1997, 272: 30611-30614]
- [Neff NF, Ellis NA, Ye TZ, Noonan J, Huang K, Sanz M, Proytcheva M. The DNA helicase activity of BLM is necessary for the correction of the genomic instability of bloom syndrome cells. Mol Biol Cell 1999, 10: 665-676]
- [Guo RB, Rigolet P, Zargarian L, Fermandjian S, Xi XG. Structural and functional characterizations reveal the importance of a zinc binding domain in Bloom's syndrome helicase. Nucleic Acids Res 2005, 33: 3109-3124]
- [Bennett RJ, Keck JL. Structure and function of RecQ DNA helicases. Crit Rev Biochem Mol Biol 2004, 39: 79-97]
- [Lee JW, Harrigan J, Opresko PL, Bohr VA. Pathways and functions of the Werner syndrome protein. Mech Ageing Dev 2005, 126: 79-86]
- [Bernstein DA, Keck JL. Conferring substrate specificity to DNA helicases: role of the RecQ HRDC domain. Structure 2005, 13: 1173-1182]
- [Wu L, Chan KL, Ralf C, Bernstein DA, Garcia PL, Bohr VA, Vindigni A, et al. The HRDC domain of BLM is required for the dissolution of double Holliday junctions. EMBO J 2005, 24: 2679-2687]
- [Mushegian AR, Bassett DE, Boguski MS, Bork P, Koonin EV. Positionally cloned human disease genes: patterns of evolutionary conservation and functional motifs. Proc Natl Acad Sci U S A 1997, 94: 5831-5836]
- [Macris MA, Krejci L, Bussen W, Shimamoto A, Sung P. Biochemical characterization of the RECQ4 protein, mutated in Rothmund-Thomson syndrome. DNA Repair (Amst) 2006, 5: 172-180]
- [Mankouri HW, Huttner D, Hickson ID. How unfinished business from S-phase affects mitosis and beyond. EMBO J 2013, 32: 2661-2671]
- [Lambert S, Carr AM. Impediments to replication fork movement: stabilisation, reactivation and genome instability. Chromosoma 2013, 122: 33-45]
- [Ralf C, Hickson ID, Wu L. The Bloom's syndrome helicase can promote the regression of a model replication fork. J Biol Chem 2006, 281: 22839-22846]
- [Machwe A, Xiao L, Groden J, Orren DK. The Werner and Bloom syndrome proteins catalyze regression of a model replication fork. Biochemistry 2006, 45: 13939-13946]
- [LeRoy G, Carroll R, Kyin S, Seki M, Cole MD. Identification of RecQL1 as a Holliday junction processing enzyme in human cell lines. Nucleic Acids Res 2005, 33: 6251-6257]
- [Karow JK, Constantinou A, Li JL, West SC, Hickson ID. The Bloom's syndrome gene product promotes branch migration of holliday junctions. Proc Natl Acad Sci U S A 2000, 97: 6504-6508]
- [Constantinou A, Tarsounas M, Karow JK, Brosh RM, Bohr VA, Hickson ID, West SC. Werner's syndrome protein (WRN) migrates Holliday junctions and co-localizes with RPA upon replication arrest. EMBO Rep 2000, 1: 80-84]
- [Boddy MN, Gaillard PH, McDonald WH, Shanahan P, Yates JR, Russell P. Mus81-Eme1 are essential components of a Holliday junction resolvase. Cell 2001, 107: 537-548]
- [Ip SC, Rass U, Blanco MG, Flynn HR, Skehel JM, West SC. Identification of Holliday junction resolvases from humans and yeast. Nature 2008, 456: 357-361]
- [Sarbajna S, West SC. Holliday junction processing enzymes as guardians of genome stability. Trends Biochem Sci 2014, 39: 409-419]
- [Bachrati CZ, Borts RH, Hickson ID. Mobile D-loops are a preferred substrate for the Bloom's syndrome helicase. Nucleic Acids Res 2006, 34: 2269-2279]
- [Suski C, Marians KJ. Resolution of converging replication forks by RecQ and topoisomerase III. Mol Cell 2008, 30: 779-789]
- [Jackson SP, Bartek J. The DNA-damage response in human biology and disease. Nature 2009, 461: 1071-1078]
- [Zou L, Elledge SJ. Sensing DNA damage through ATRIP recognition of RPA-ssDNA complexes. Science 2003, 300: 1542-1548]
- [Esashi F, Christ N, Gannon J, Liu Y, Hunt T, Jasin M, West SC. CDK-dependent phosphorylation of BRCA2 as a regulatory mechanism for recombinational repair. Nature 2005, 434: 598-604]
- [Ceballos SJ, Heyer WD. Functions of the Snf2/Swi2 family Rad54 motor protein in homologous recombination. Biochim Biophys Acta 2011, 1809: 509-523]
- [Nimonkar AV, Ozsoy AZ, Genschel J, Modrich P, Kowalczykowski SC. Human exonuclease 1 and BLM helicase interact to resect DNA and initiate DNA repair. Proc Natl Acad Sci U S A 2008, 105: 16906-16911]
- [Wu L, Davies SL, Levitt NC, Hickson ID. Potential role for the BLM helicase in recombinational repair via a conserved interaction with RAD51. J Biol Chem 2001, 276: 19375-19381]
- [Hu Y, Raynard S, Sehorn MG, Lu X, Bussen W, Zheng L, Stark JM, et al. RECQL5/Recql5 helicase regulates homologous recombination and suppresses tumor formation via disruption of Rad51 presynaptic filaments. Genes Dev 2007, 21: 3073-3084]