Paměťové B lymfocyty
Paměťové B lymfocyty jsou B buňky, které dlouhodobě přežívají po kontrakci primární imunitní odpovědi a zprostředkovávají imunitní paměť. Při opakovaném setkání s antigenem reagují rychleji a intenzivněji, produkují obvykle přesmyknuté vysokoafinní protilátky, a na jejich vlastnostech je založena účinnost vakcín[1]. Protilátky mohou indukovat aktivaci komplementu, cytotoxických buněk, opsonizovat a neutralizovat. Jsou mocnou zbraní proti celé řadě infekčních onemocnění. Protilátkovou paměť zajišťují vedle paměťových B buněk také dlouho-žijící plasmatocyty, jejichž protilátky tvoří první obrannou linii[2]. Jsou-li přítomny v dostatečném množství, neutralizují patogena při vstupu do organismu a k aktivaci paměťových B buněk nedojde.
.jpg.webp)
Lidské paměťové B buňky se nachází zejména v sekundárních lymfatických orgánech, tvoří téměř polovinu všech B buněk sleziny. Můžeme je nalézt také v krvi, ovšem v menším množství[2]. Většina poznatků o paměťových B lymfocytech je však odvozena ze studií na myších modelech a není proto jisté, že vše funguje u lidí stejně[1].
Typy paměťových B lymfocytů
Paměťové B lymfocyty dělíme podle způsobu jejich vzniku na paměťové B lymfocyty z germinálních center a paměťové B lymfocyty na germinálních centrech nezávislé. Dále rozlišujeme T - závislé a T - nezávislé paměťové B lymfocyty. Rovněž je lze dělit podle typu BCR, který nesou na svém povrchu, na IgM+, IgG+ a IgA+[1].
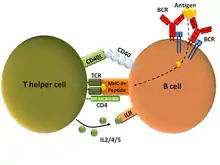
T - závislé B lymfocyty
Taktéž označovány jako B2 buňky. Při prvním setkání s antigenem (tzv. primární imunitní odpověď) se aktivují díky vazbě antigenu na jejich BCR a pomoci od folikulárních T lymfocytů (Tfh). K interakci B a T buněk dochází v rámci sekundárních lymfatických orgánů, jako jsou třeba lymfatické uzliny, na rozhraní tzv. T buněčné zóny a B folikulu. Aktivovaná B buňka proliferuje, může provádět izotypový přesmyk, a její klonální potomci mohou diferencovat v krátko-žijící plasmatické buňky, paměťové B buňky a nebo vytvářet germinální centra. V rámci germinálních center dochází k afinitní maturaci (somatické hypermutaci) B buněk. Výsledkem jsou dlouho-žijící plasmatické buňky a opět paměťové B buňky[1]. Ve který podtyp B buňka diferencuje je určováno řadou faktorů, mezi nimi například mírou afinity BCR a antigenu, též cytokiny[3].
Paměťové B lymfocyty na germinálních centrech nezávislé
Jedná se o paměťové B buňky, které diferencují z aktivovaných B lymfocytů před vstupem do germinálního centra. Jejich BCR má relativně nízkou afinitu k antigenu a interakce s Tfh je slabší. To proto, že B buňky interagují s Tfh prostřednictvím MHCII glykoproteinů. Čím nižší má BCR afinitu k antigenu, tím méně antigenů B buňka vychytá, pohltí a vystaví na svém MHCII pro Tfh. Následně neobdrží dostatečně intenzivní a dlouhotrvající signál, v podobě interakce CD40-CD40L a cytokinů, a proto nevytvoří germinální centrum[3][1]. Tyto B buňky mohou provést izotypový přesmyk, neprocházejí však somatickou hypermutací, jejich afinita k antigenu zůstává relativně slabá. Tělo si tak v jejich podobě udržuje širokou škálu paměťových B buněk, které mohou reagovat i proti patogenům, kteří jsou pouze příbuzní těm, se kterými se tělo již setkalo[1][4].
Paměťové B lymfocyty z germinálních center
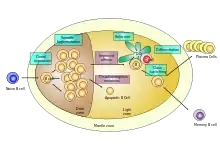
Germinální centrum je transientní útvar v B folikulu tvořený aktivovanými dělícími se B lymfocyty, Tfh lymfocyty a folikulárními dendritickými buňkami (FDC). Rozlišujeme v rámci něj tmavou a světlou zónu. V tmavé zóně se B lymfocyty rychle dělí a dochází u nich k somatické hypermutaci. Ta spočívá v indukci náhodných bodových mutací v oblasti variabilních úseků imunoglobulinových řetězců. S určitou pravděpodobností vznikají takové mutace, které zvýší afinitu protilátky k antigenu. B lymfocyt s takto vylepšeným BCR (což je membránově vázaná varianta protilátky) je následně vyselektován pomocí Tfh v oblasti světlé zóny. Ve světlé zóně jsou přítomny Tfh a FDC. FDC na svém povrchu zachytávají velké množství antigenů a nabízejí je mutovaným B lymfocytům k interakci. B buňka s vysokoafinním BCR zachytí velké množství příslušných antigenů, pohltí je, vystaví na MHCII a interaguje s Tfh intenzivně. V závislosti na intenzitě interakce s Tfh (a dalších faktorech) může B buňka absolvovat hypermutační okruh ještě jednou, nebo germinální centrum opouští a diferencuje v dlouho-žijící plasmatické buňky či v paměťové B buňky z germinálních center. B buňky, které obdržely nejvíce signálů od Tfh, v opakovaných kolech germinální reakce proliferují intenzivněji, než ostatní. Převáží tak B buňky nesoucí vysokoafinní BCR[5][1]. Výsledkem germinální reakce jsou pak plazmatické a paměťové B buňky schopné produkce vysokoafinních protilátek, proto bývá tento proces označován jako afinitní maturace.
Doposud není přesně známo, na základě jakého mechanismu vznikají z afinitně maturovaných B lymfocytů paměťové B buňky. Zvažována je existence určitého významného transkripčního regulátoru. S indukcí jeho exprese by došlo ke spuštění diferenciačního programu v paměťovou B buňku. Zatím se však nezdařilo takový transkripční faktor identifikovat. Druhou možností je, že paměťové B buňky vznikají z těch B lymfocytů, které jsou schopné přežívat nejdéle. Od zbytku se odlišují expresí některých anti-apoptotických proteinů (Bcl-2), které jim přináší výhodu v přežívání, či potlačením exprese pro-apoptotických genů (Bim). A pravděpodobně zde bude hrát roli opět afinita BCR a míra interakce s Tfh v germinálních centrech[1][3].
IgM+ paměťové B lymfocyty
Na svém povrchu nesou nepřesmyknutý BCR, v podobě IgM - tzn. zatím u nich k izotypovému přesmyku nedošlo. Jsou podobné naivním B buňkám. Po opětovném setkání s antigenem hodně proliferují a často vstupují do germinální reakce[1].
IgG+ a IgA+ paměťové B lymfocyty
Na svém povrchu nesou přesmyknutý BCR, v podobě IgG či IgA, izotypový přesmyk u nich už proběhl. IgG+ po opětovném setkání s antigenem typicky diferencují v plasmatické buňky a proliferují méně[1].
Pro přežívání paměťových buněk, které prošly izotypovým přesmykem, jsou významné transkripční faktory spojené s jeho indukcí (například T-bet pro IgG či RORα pro IgA)[1].
IgE+ paměťové B lymfocyty
Zatím se nezdařilo je in vivo detekovat. Pokud existují, tak ve velmi malých počtech. Pravděpodobnější se zdá varianta, že paměťové IgE protilátky jsou produkovány IgG+ paměťovými B lymfocyty, které prošly afinitní maturací před svým vznikem, a po opětovném setkání s antigenem u nich proběhne znovu izotypový přesmyk[1]. Začnou pak produkovat vysokoafinní IgE, které je podstatou alergických onemocnění[6].
Vlastnosti paměťových B lymfocytů
Typickými vlastnostmi paměťových B lymfocytů je schopnost dlouhodobě, u člověka po desítky let[2], přežívat v organismu v quiescentním stavu, po opětovné interakci s antigenem se rychle reaktivovat, dělit, dát vznik plazmatickým buňkám i vstupovat znovu do germinální reakce a dále zvyšovat afinitu produkovaných protilátek. Každá sekundární imunitní odpověď na daný antigen je tak intenzivnější než odpověď primární[7][1].
Schopnost namnožit se a dát vznik celé populaci specifických B lymfocytů se někdy označuje jako kmenovost paměťových B lymfocytů. Zdá se, že IgM+ paměťové B lymfocyty jsou v tomto směru obzvláště schopné[1].
Dlouhodobé přežívání je spojeno zejména se změnami metabolismu a blokováním apoptózy. Nezbytná je přítomnost FDC a tonická signalizace přes BCR, která indukuje expresi anti-apoptotických genů. Antigen jako takový přítomen být nemusí, ani T lymfocyty nejsou k přežívání paměťových B buněk nezbytné. Myší IgM+ paměťové B buňky přežívají lépe než IgG+, u lidských rozdíl nejspíše není[1].
Rychlá a intenzivní odpověď paměťových B buněk oproti naivním B buňkám může být částečně důsledkem přesmyknutého BCR. Izotypový přesmyk mění Fc část protilátky, která v případě BCR zasahuje dovnitř buňky a může se podílet na signalizaci. Bylo to studováno na IgG1 typu BCR, který má oproti IgM větší konzervovanou cytoplazmatickou doménu, která interaguje s komponentami MAP-kinázových kaskád. Kromě toho hraje významnou roli stimulační historie paměťového B lymfocytu, v důsledku níž má pozměněné hladiny transkripčních faktorů[1].
Reaktivace paměťových B lymfocytů je závislá na interakci s paměťovými Tfh lymfocyty skrze MHCII. Oba typy paměťových buněk se nachází v sekundárních lymfatických tkáních v oblasti B folikulu a mohou spolu rychle interagovat, pakliže se objeví antigen, ke kterému jsou oba specifičtí. Interakce je zřejmě oboustranná, v první fázi B buňka funguje jako antigen-prezentující buňka a aktivuje Tfh. Aktivovaný Tfh pak aktivuje paměťovou B buňku. Významnou úlohu sehrávají také FDC, které rychle vychytávají přicházející antigeny, a umožňují B lymfocytům s nimi interagovat. Zejména pokud jsou antigeny zachyceny dříve produkovanými protilátkami[1].
Opětovný vstup do germinální reakce je typický hlavně pro IgM+ paměťové B buňky. Germinální centra v některých případech nevymizí po primární imunitní reakci a reaktivované paměťové B buňky se mohou znovu připojit. Mohou také procházet izotypovým přesmykem a dát vznik plasmatickým buňkám produkujícím vysokoafinní IgG protilátky. Na zvyšování afinity produkovaných protilátek se podílí také proces reaktivace, kdy interakce s Tfh vede k selekci B buněk s vyšší afinitou BCR[1].
T - nezávislé B lymfocyty
Taktéž označovány jako B1 buňky. Pro aktivaci a následnou produkci protilátek nevyžadují pomoc od T lymfocytů. Produkují přirozené IgM protilátky po rozpoznání zejm. bakteriálních multivalentních antigenů (sacharidové struktury). Ty "prováží" jejich BCR natolik, že indukovaný signál je dostatečný sám o sobě k aktivaci B lymfocytu. Nachází se hlavně v peritoneální a pleurální dutině, minoritně ve slezině. Některé rozpoznávají též naše vlastní antigeny[8].
Existence paměťových B1 buněk byla dokázána teprve nedávno. Jsou udržovány v peritoneální dutině a po opětovném setkání s antigenem se aktivují. Na rozdíl od klasických B2 paměťových buněk, zřejmě nemají žádné speciální vlastnosti, které by následně umožňovaly rychlejší a intenzivnější imunitní odpověď. Paměť je zde dána pouze udržením většího počtu buněk specifických k danému antigenu[1].
Povrchové markery paměťových B lymfocytů
Typickým markerem lidských paměťových B lymfocytů je CD27. Dále obvykle exprimují na povrchu přesmyknuté BCR, tedy jiného než IgM či IgD typu[2][7].
Reference
- KUROSAKI, Tomohiro; KOMETANI, Kohei; ISE, Wataru. Memory B cells. Nature Reviews Immunology. 2015/03, roč. 15, čís. 3, s. 149–159. Dostupné online [cit. 2018-02-05]. ISSN 1474-1741. DOI 10.1038/nri3802. (En)
- HAUSER, Anja E.; HÖPKEN, Uta E. B Cell Localization and Migration in Health and Disease. [s.l.]: [s.n.] Dostupné online. DOI 10.1016/b978-0-12-397933-9.00012-6. S. 187–214.
- SHINNAKASU, Ryo; KUROSAKI, Tomohiro. Regulation of memory B and plasma cell differentiation. Current Opinion in Immunology. Roč. 45, s. 126–131. Dostupné online [cit. 2018-02-06]. DOI 10.1016/j.coi.2017.03.003.
- PUPOVAC, Aleta; GOOD-JACOBSON, Kim L. An antigen to remember: regulation of B cell memory in health and disease. Current Opinion in Immunology. Roč. 45, s. 89–96. Dostupné online [cit. 2018-02-06]. DOI 10.1016/j.coi.2017.03.004.
- DE SILVA, Nilushi S.; KLEIN, Ulf. Dynamics of B cells in germinal centres. Nature Reviews Immunology. 2015/03, roč. 15, čís. 3, s. 137–148. Dostupné online [cit. 2018-02-06]. ISSN 1474-1741. DOI 10.1038/nri3804. (En)
- GOULD, Hannah J.; SUTTON, Brian J. IgE in allergy and asthma today. Nature Reviews Immunology. 2008/03, roč. 8, čís. 3, s. 205–217. Dostupné online [cit. 2018-02-06]. ISSN 1474-1741. DOI 10.1038/nri2273. (En)
- M.),, Murphy, Kenneth (Kenneth. Janeway's immunobiology. Ninth edition. vyd. New York, NY, USA: [s.n.] xx, 904 pages s. ISBN 9780815345510. OCLC 933586700
- MONTECINO-RODRIGUEZ, Encarnacion; DORSHKIND, Kenneth. B-1 B Cell Development in the Fetus and Adult. Immunity. Roč. 36, čís. 1, s. 13–21. Dostupné online [cit. 2018-02-06]. DOI 10.1016/j.immuni.2011.11.017.