Hadrosauridae
Hadrosauridi (čeleď Hadrosauridae) byli velmi rozšířenou čeledí ptakopánvých dinosaurů, tzv. kachnozobých dinosaurů. Patřili do nadčeledi Hadrosauroidea a představovali dominantní formy býložravců v mnoha pozdně křídových ekosystémech severních kontinentů. Jméno dal této skupině první rozeznaný taxon, Hadrosaurus foulkii, popsaný roku 1858 z východu USA.[1]
Stratigrafický výskyt: Svrchní křída, asi před 86 až 66 miliony let | |
|---|---|
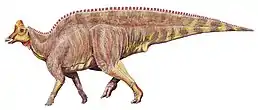 Magnapaulia laticaudus, zástupce čeledi | |
| Vědecká klasifikace | |
| Říše | živočichové (Animalia) |
| Kmen | strunatci (Chordata) |
| Třída | plazi (Sauropsida) |
| Nadřád | dinosauři (Dinosauria) |
| Řád | ptakopánví (Ornithischia) |
| Podřád | Ornithopoda |
| Nadčeleď | Hadrosauroidea |
| Čeleď | Hadrosauridae Cope, 1869 |
| Některá data mohou pocházet z datové položky. | |

Popis a význam
Žili především v pozdní křídě a hráli roli dominantních býložravců tohoto posledního období existence dinosaurů. Jejich pozůstatky jsou dnes známy z Asie, Evropy i Severní Ameriky. Patří mezi vývojově vyspělé ptakopánvé dinosaury a jejich předky byli dinosauři z příbuzenstva rodu Iguanodon. Tato čeleď se dělí do dvou podčeledí - Hadrosaurinae a Lambeosaurinae (kteří vykazují přítomnost hlavových útvarů vzhledu hřebenů nebo trubic). Největší zástupci této skupiny, jako čínský rod Shantungosaurus nebo severoamerický Lambeosaurus mohli dosahovat délky až 17 metrů a hmotnosti přes 10 (možná však až kolem 20) tun.[2] Dinosauři z této skupiny patří mezi nejlépe dochované zkameněliny této skupiny živočichů vůbec.
Jejich geografické rozšíření v období pozdní křídy bylo značné a zahrnovalo velkou část Severní Ameriky, Evropy i Asie.[3] Ačkoliv se jednalo ve velké většině případů o obyvatele severní polokoule, existují také fosilie druhů, obývajících pevniny jižního superkontinentu Gondwany. Zejména se jednalo o argentinské rody Secernosaurus, Willinakaqe a Bonapartesaurus. Velmi rozšířenou skupinou byli například lambeosaurini, známí z území několika kontinentů.[4]
Dinosauří mumie
Tzv. "dinosauří mumie" patří právě hadrosauridům, především rodům Edmontosaurus a Brachylophosaurus. U skvěle zachovaných jedinců, známých jako "Leonardo" (stáří 77 milionů let) a "Dakota" (67 milionů let) byly objeveny i obrysy vnitřních orgánů a svalstva nebo otisky kůže. Výjimečně se mohou dochovat i původní proteiny ve fosiliích těchto dinosaurů (např. u exempláře brachylofosaura s označením MOR 2598).[5] V některých případech je dochována textura kůže v trojrozměrném provedení a umožňuje také podrobný histologický výzkum.[6]
Neobvykle dobře dochovaný fosilní otisk kůže hadrosauridního dinosaura byl předběžně formálně popsán také koncem roku 2019. Exemplář s katalogovým označením YPMPU 016969 představuje otisk z boku (trupu) hadrosaurida a poskytuje cenné informace o stavbě a tloušťce kůže hadrosauridů i o jejím někdejším zbarvení a struktuře.[7]
Paleobiologie
Původně se paleontologové domnívali, že hadrosauridi byli výbornými plavci, kteří se ve vodě poháněli pohyby mohutného ocasu a k plavání jim sloužila i prstová blána, o jejíž přítomnosti zdánlivě svědčily některé fosilie. Dnes předpokládáme, že kachnozobí dinosauři byli téměř výlučně po suché zemi se pohybující obratlovci.[8] V roce 2008 bylo novým podrobným výzkumem zjištěno, že hadrosauridi z podčeledi Lambeosaurinae se pravděpodobně dorozumívali táhlými bučivými zvuky, které vydávali pomocí výrazných "hřebenů" na hlavě. Ty sloužily patrně jako rezonanční komory pro zesílení intenzity táhlých bučivých zvuků.[9][10]
Bylo také zjištěno, že tito dinosauři velmi rychle rostli, neboť z původní délky kolem 0,8 metru po vylíhnutí dosahovali dospělé délky kolem 9 a více metrů již během 7. nebo 8. roku života. Rostli tedy podstatně rychleji než například draví teropodní dinosauři tyranosauridi.[11] Jak ukázaly objevy sérií stop, někteří hadrosauridi pochodovali v "rodinných" stádech s různě starými (a tedy velkými) jedinci. Přečkávali také polární zimu. [12] Velkou úlohu v evolučním úspěchu této skupiny zřejmě sehrály zubní baterie hadrosauridů, umožňující velmi efektivní mechanické zpracování tuhé rostlinné potravy.[13] V roce 2017 byla publikována vědecká studie, podle které hadrosauridi na území dnešního Utahu v době před 75 miliony let pojídali s rozkládající se dřevní hmotou i velké suchozemské korýše, přičemž jejich pozření jim snad mohlo doplňovat vápník a další potřebné látky.[14][15] Dentální baterie hadrosauridů byly do značné míry unikátní a vykazovaly jistý stupeň dynamického pohybu.[16][17] Navzdory běžně tradovanému názoru se ramfotéka hadrosauridů nepodobala kachnímu zobáku a jejich označení "kachnozobí" dinosauři tak není příliš přesné.[18] Objev ze Španělska ukázal, že hadrosauridi žili ve skupinách s převažujícím množstvím mláďat a menším počtem plně dospělých jedinců.[19]
Výzkum histologie fosilních kostí mongolských hadrosauridů dokazuje, že u obřích druhů zřejmě evolucí došlo k nahrazení přerušovaného růstu kontinuálním, který vedl k dosažení obřích rozměrů u rodů, jako je Shantungosaurus, Edmontosaurus, Saurolophus a dalších.[20]
V roce 2020 byla publikována studie o identifikaci histiocytózy Langerhansových komůrek ve fosilních kostech hadrosaurida, formy benigního kostního karcinomu, známého i u člověka.[21] Objev fosilních obratlů s vetknutými fragmenty tyranosauřích zubů odhalil, že hadrosauridi pravděpodobně neměli meziobratlové ploténky.[22] Kachnozobí dinosauři obecně trpěli relativně vysokým výskytem kostních tumorů, které jsou jako jediné dobře zjistitelné i z fosilního záznamu.[23]
Zkamenělá vajíčka hadrosauridů jsou známá již dlouhou dobu. Patří mezi ně také oorod Stromatoolithus.[24]
Migrace a stádní chování
Výzkum chemického složení zubů hadrosauridů z Utahu naznačuje, že stádní chování hadrosauridů do značné míry ovlivňovaly sezónní záplavy. Populace vzdálené od sebe pouze 5 až 15 kilometrů tak pravděpodobně využívaly jiné zdroje potravy ve své ekologické nice.[25] Obvykle tito dinosauři migrovali na vzdálenost maximálně kolem 80 kilometrů, nikoliv na celé tisíce kilometrů, jak bylo dříve některými badateli předpokládáno. Ukázal to výzkum izotopů stroncia u fosilií kachnozobých dinosaurů v kanadské Albertě.[26]
Zástupci skupiny
- Čeleď Hadrosauridae
- Podčeleď Hadrosaurinae
- Podčeleď Saurolophinae
- Podčeleď Lambeosaurinae
Odkazy
Reference
- http://www.pravek.info/zajimavosti/pribeh-prvniho-severoamerickeho-dinosaura/
- Xing, Hai; Zhao, Xijin; Wang, Kebai; Li, Dunjing; Chen, Shuqing; Mallon, Jordan C; Zhang, Yanxia; Xu, Xing (2014). "Comparative osteology and phylogenetic relationship of Edmontosaurus and Shantungosaurus (Dinosauria: Hadrosauridae) from the Upper Cretaceous of North America and East Asia". Acta Geologica Sinica-English Edition. 88 (6): 1623–1652.
- David G. Taylor and Spencer G. Lucas (2018). A Late Cretaceous (Campanian) hadrosaur sacrum from the Cape Sebastian Sandstone, Curry County, Oregon. Lucas, S.G. and Sullivan, R.M., eds., 2018, Fossil Record 6. New Mexico Museum of Natural History and Science. 79: 695-702.
- SOCHA, Vladimír. Hřeben plný slov. OSEL.cz [online]. 20. května 2021. Dostupné online. (česky)
- http://dinosaurusblog.com/2017/01/30/kolagen-stary-78-milionu-let/
- Mauricio Barbi, Phil R. Bell, Federico Fanti, James J. Dynes, Anezka Kolaceke, Josef Buttigieg & Philip J. Currie (2019). Integumentary structure and composition in an exceptionally well-preserved hadrosaur (Dinosauria: Ornithischia). PeerJ Preprints 7:e27698v1. doi: https://doi.org/10.7287/peerj.preprints.27698v1
- Matteo Fabbri, Jasmina Wiemann, Fabio Manucci & Derek E. G. Briggs (2019). Three‐dimensional soft tissue preservation revealed in the skin of a non‐avian dinosaur. Palaeontology (advance online publication). doi: https://doi.org/10.1111/pala.12470
- Bakker, R. T. (1986). The Dinosaur Heresies, Zebra Books, str. 148.
- Archivovaná kopie. dinosaurusblog.com [online]. [cit. 2018-04-18]. Dostupné v archivu pořízeném dne 2018-07-22.
- SOCHA, Vladimír. Sto let dinosaura s lebeční trubicí. OSEL.cz [online]. 8. dubna 2022. Dostupné online. (česky)
- Archivovaná kopie. dinosaurusblog.com [online]. [cit. 2018-04-18]. Dostupné v archivu pořízeném dne 2018-06-23.
- Archivovaná kopie. dinosaurusblog.com [online]. [cit. 2018-04-18]. Dostupné v archivu pořízeném dne 2018-06-08.
- http://dinosaurusblog.com/2016/10/03/zubni-baterie-hadrosauridu/
- SOCHA, Vladimír. Kachnozobí dinosauři pojídali korýše. OSEL.cz [online]. 23. října 2017. Dostupné online. (česky)
- Karen Chin, Rodney M. Feldmann, Jessica N. Tashman (2017). Consumption of crustaceans by megaherbivorous dinosaurs: dietary flexibility and dinosaur life history strategies. Scientific Reports, 7 (1): 11163; doi: 10.1038/s41598-017-11538-w
- Katherine Bramble, Aaron R. H. LeBlanc, Denis O. Lamoureux, Mateusz Wosik & Philip J. Currie (2017). Histological evidence for a dynamic dental battery in hadrosaurid dinosaurs. Scientific Reports 7, Article number: 15787 (2017) doi:10.1038/s41598-017-16056-3
- https://aaronrhleblanc.wordpress.com/2018/11/08/dinosaur-dentistry-part-3-steak-knives-and-dental-batteries/
- https://blogs.scientificamerican.com/tetrapod-zoology/enough-with-the-duckbilled-dinosaurs/#
- Víctor Fondevilla; et al. (2018). Ontogeny and taxonomy of the hadrosaur (Dinosauria, Ornithopoda) remains from Basturs Poble bonebed (late early Maastrichtian, Tremp Syncline, Spain). PLoS ONE 13(10): e0206287. doi: https://doi.org/10.1371/journal.pone.0206287
- Justyna Słowiak, Tomasz Szczygielski, Michał Ginter & Łucja Fostowicz‐Frelik (2020). Uninterrupted growth in a non‐polar hadrosaur explains the gigantism among duck‐billed dinosaurs. Palaeontology. doi: https://doi.org/10.1111/pala.12473
- Bruce M. Rothschild, Darren Tanke, Frank Rühli, Ariel Pokhojaev & Hila May (2020). Suggested Case of Langerhans Cell Histiocytosis in a Cretaceous dinosaur. Scientific Reports, 10, Article number: 2203. doi: https://doi.org/10.1038/s41598-020-59192-z
- Bruce M. Rothschild, Robert A. Depalma, David A. Burnham & Larry Martin (2020). Anatomy of a dinosaur--Clarification of vertebrae in vertebrate anatomy. Anatomia, Histologia, Embryologia. doi: https://doi.org/10.1111/ahe.12573
- Rozhovor s V. Sochou o nemocech dinosaurů v rozhlasovém pořadu Meteor, čas 19:55 min. (vysíláno 30. 10. 2021)
- Xufeng Zhu, Qiang Wang & Xiaolin Wang (2021). Restudy of the original and new materials of Stromatoolithus pinglingensis and discussion on some Spheroolithidae eggs. Historical Biology (advance online publication). doi: https://doi.org/10.1080/08912963.2021.1910817
- Victoria F. Crystal, Erica S. J. Evans, Henry Fricke, Ian M. Miller & Joseph J. W. Sertich (2018). Late Cretaceous fluvial hydrology and dinosaur behavior in southern Utah, USA: Insights from stable isotopes of biogenic carbonate. Palaeogeography, Palaeoclimatology, Palaeoecology. doi: https://doi.org/10.1016/j.palaeo.2018.11.022
- David F. Terrill, Charles M. Henderson and Jason S. Anderson (2020). New application of strontium isotopes reveals evidence of limited migratory behaviour in Late Cretaceous hadrosaurs. Biology Letters, 16(3): 20190930. doi: https://doi.org/10.1098/rsbl.2019.0930
Literatura
- Thomas L. Stubbs, Michael J. Benton, Armin Elsler and Albert Prieto-Márquez (2019). Morphological innovation and the evolution of hadrosaurid dinosaurs. Paleobiology 45(2): 347-362. doi: https://doi.org/10.1017/pab.2019.9
- Héctor E. Rivera-Sylva; et al. (2019). Preliminary assessment of hadrosaur dental microwear from the Cerro del Pueblo Formation (Upper Cretaceous: Campanian) of Coahuila, northeastern Mexico. Paleontología Mexicana. 8 (1): 17-28.
- Mateusz Wosik, Kentaro Chiba, François Therrien and David C. Evans (2020). Testing size--frequency distributions as a method of ontogenetic aging: a life-history assessment of hadrosaurid dinosaurs from the Dinosaur Park Formation of Alberta, Canada, with implications for hadrosaurid paleoecology. Paleobiology. doi: https://doi.org/10.1017/pab.2020.2
- Taia Wyenberg-Henzler, R. Timothy Patterson & Jordan C. Mallon (2022). Ontogenetic dietary shifts in North American hadrosaurids. Cretaceous Research. 105177. doi: https://doi.org/10.1016/j.cretres.2022.105177
Externí odkazy
 Obrázky, zvuky či videa k tématu Hadrosauridae na Wikimedia Commons
Obrázky, zvuky či videa k tématu Hadrosauridae na Wikimedia Commons - Článek o potravních návycích hadrosauridů (anglicky)
- Článek o rozmnožování hadrosauridů (anglicky)
- SOCHA, Vladimír. Křídový karibu. OSEL.cz [online]. 22. června 2020. Dostupné online. (česky)