Tyrosinkinázový receptor
Tyrozinkinázový receptor (anglicky RTK - receptor tyrosine kinase) je označení pro skupinu receptorů, které se nacházejí na vnějším povrchu membrán buněk. Obsahují tyrosinkinázu, enzym, který přenáší fosfátovou skupinu na aminokyselinu tyrosin. RTK mají vysokou afinitu k polypeptidům, jako jsou růstové faktory, cytokiny a některé hormony (především inzulin). Tyrosinkinasové receptory mají důležitou úlohu v rozvoji a vývoji mnoha typů rakoviny.[1]
V lidském genomu bylo objeveno přes 90 genů pro tyrosinkinázu. Z toho 58 genů obsahuje genetický kód pro bílkoviny tyrosinkinásového receptoru.[2]
Struktura
Mnoho tyrosinkinásových receptorů jsou monomery (jejich velká molekula tvoří jedinou podjednotku), ale některé další, jako např. inzulínový receptor, jsou oligomery (receptor se skládá z dvou či více podjednotek). V případě inzulínového receptoru se konkrétně jedná o dimer. Každá podjednotka má transmembránovou proteinovou doménu složenou z řetězce 25-38 aminokyselin, jehož N-konec je na extracelulárním konci (u inzulínového receptoru se označuje jako α) a C-konec na intracelulárním (β). Extracelulární N-konec řetězce je složen z velké proteinové domény, ke které se připojují výše zmíněné ligandy. Mimobuněčný C-konec řetězce obsahuje tyrosinkinasové domény zodpovědné za tyrosinkinasové aktivitu těchto receptorů.
Kinázová aktivita
V biochemii je kináza typem enzymu, který přenáší fosfátové skupiny z molekul s makroergními vazbami (především z ATP) na specifickém cílové molekuly, čímž je povyšuje na vyšší energetický stav a ony díky tomu změní svou konformaci, což vede ke změně biologické aktivity takto fosforylované cílové molekuly. Celý proces se nazývá fosforylace. Enzym, který provádí opačnou reakci, se nazývá fosfatáza. Kinázy, které specificky v daném bílkovinném řetězci specificky fosforylují aminokyselinu tyrosin, se nazývají právě tyrosinkinasy.
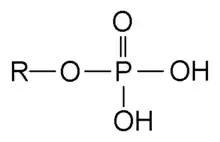
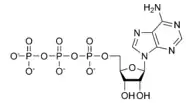
Navázání růstového faktoru na extracelulární doménu RTK spustí dimerizaci se sousedním receptorem stejného druhu. (Dimerizace je spojení dvou monomerů). Dimerizace vede k rychlé aktivaci cytoplazmatických domén tyrosinkinasy, která následně autofosforyluje tyrosinové zbytky v intracelulárních doménách receptoru, čímž dojde k celkové aktivaci receptoru.
Transdukce signálu
Fosforylované specifické tyrosinové zbytky společně s celým aktivovaným receptorem vytváří vazebná místa pro SH2 domény a PTB domény.[3] Specifické proteiny, které obsahují tyto domény, jsou Src a fosfolipáza C. Fosforylace a aktivace těchto dvou proteinů je počátečním krokem v transdukci signálu. Jiné proteiny, které se navazují na aktivovaný receptor působí jako tzv. adaptorové proteiny. Samy o sobě nemají enzymovou aktivitu. Tyto adaptorové proteiny představují začátek jiných alternativních signálních drah, jako např. signální kaskády MAP kinasy.[1]
Třídy tyrosinkinázových receptorů
Bylo objeveno přibližně 20 tříd. Některé z nich nesou tyto anglické názvy:[4]
- RTK class I (EGF receptor family)
- RTK class II (Insulin receptor family)
- RTK class III (Platelet-derived growth factor receptor|PDGF receptor family)
- RTK class IV (Fibroblast growth factor receptor z FGF receptor family)
- RTK class V (VEGF receptors family)
- RTK class VI (C-Met|HGF receptor family)
- RTK class VII (Trk receptor family)
- RTK class IX (AXL receptor family)
- RTK class X (LTK receptor family)
- RTK class XI (Angiopoietin Receptors: Tie-1 & Tie-2 z TIE receptor family)
- RTK class XII (ROR receptor family)
- RTK class XIII (DDR receptor family)
- RTK class XV (KLG receptor family)
- RTK class XVI (RYK receptor family)
- RTK class XVII (MuSK receptor family)
Rodiny
Rodina receptorů pro FGF
Fibroblastové růstové faktory (FGF - Fibroblast growth factor receptor) jsou největší rodinou růstových faktorů sestávající z 23 členů.[5] Při přirozeném posttranslačním sestřihu týkajícího se čtyř genů pro FGFR dochází k tvorbě 48 odlišných izoforem FGFR.[6] Tyto izoformy se odlišují ve vlastnostech svých vazebných a kinasových domén, ačkoliv všechny obsahují shodnou extracelulární oblast složenou z tří imunoglobinu-podobných (Ig) domén (D1-D3), čímž náleží do nadrodiny imunoglobinů.[7] K interakcím s FGF dochází cestou FGFR domén D2 a D3. Každý receptor může být aktivován různými fibroblastovými růstovými hormony. FGF mohou taktéž aktivovat více než jen jeden druh receptoru, což však neplatí pro FGF-7, který může aktivovat pouze FGFR2b.[6] Gen pro pátý FGFR protein, FGFR5, byl také objeven. Na rozdíl od FGFR 1-4 postrádá cytoplasmatickou tyrosinkinasovou doménu a jediná isoforma tohoto receptoru obsahuje pouze extracelulární domény D1 a D2.[8]
Rodina receptorů pro VEGF
Vaskulární endotelový růstový faktor (VEGF - Vascular endothelial growth factor) významně přispívá k proliferaci endoteliálních buněk cév a ke zvýšení celkové permeability cév. Dva specifické tyrosinkinasové receptory dokáží vázat VEGF: VEGFR-1 a VEGFR-2.[9]
VEGF receptory mají extracelulární část sestávající ze sedmi imunoglobinu-podobných domén, proto jako v případě receptorů pro FGF patří do nadrodiny imunuglobinů. Také obsahují jednu transmembránovou oblast a jednu intracelulární oblast obsahující roztroušené tyrosinkinasové domény. VEGF-A se váže na VEGFR-1 a VEGFR-2. VEGFR-2 má mnohem větší význam pro signalizaci vaskulárního endotelového růstového faktoru než VEGFR-1, jehož funkce je méně známa. Předpokládá se, že VGFR-1 má určitou úlohu v modulaci signálu z VEGFR-2. Jiná funkce VEGFR-1 může být úloha jako "odlákávajícího" receptoru, který snižuje množství ligandů vázaných na FEGFR-2. Toto se zdá být nejdůležitější v průběhu vaskulogeneze u embrya. Třetí receptor byl také objeven (VEGFR-3). VEGF-A nemá k němu prakticky žádnou afinitu. VEGFR-3 je důležitý v lymfangiogenezi.
Reference
V tomto článku byl použit překlad textu z článku Receptor tyrosine kinase na anglické Wikipedii.
- Zwick, E. Bange, J. Ullrich, A. Receptor tyrosine kinase signalling as a target for cancer intervention strategies. Endocr. Relat. Cancer. 2001, roč. 8, čís. 3, s. 161–173. DOI 10.1677/erc.0.0080161. PMID 11566607.
- Robinson DR, Wu YM, Lin SF. The protein tyrosine kinase family of the human genome. Oncogene. 2000, roč. 19, čís. 49, s. 5548–5557. DOI 10.1038/sj.onc.1203957. PMID 11114734.
- Pawson, T. Protein modules and signalling networks. Nature. 1995, roč. 373, čís. 6515, s. 573–580. DOI 10.1038/373573a0. PMID 7531822.
- www.genome.ad.jp Retrieved on 2007-04-05
- Ornitz DM. and Itoh, N. Fibroblast growth factors. Genome Biol.. 2001, roč. 2, čís. 3, s. REVIEWS 3005. DOI 10.1186/gb-2001-2-3-reviews3005. PMID 11276432.
- Duchesne L, Tissot B. et al. N-glycosylation of fibroblast growth factor receptor 1 regulates ligand and heparan sulfate co-receptor binding. J. Biol. Chem.. 2006, roč. 281, čís. 37, s. 27178–27189. DOI 10.1074/jbc.M601248200. PMID 16829530.
- Coutts JC, and Gallagher JT. Receptors for fibroblast growth factors. Immunol. Cell. Biol.. 1995, roč. 73, čís. 6, s. 584–589. DOI 10.1038/icb.1995.92. PMID 8713482.
- Sleeman M, Fraser J. et al. Identification of a new fibroblast growth factor receptor, FGFR5. Gene. 2001, roč. 271, čís. 2, s. 171–182. DOI 10.1016/S0378-1119(01)00518-2. PMID 11418238.
- Robinson, CJ and Stringer, SE. The splice variants of vascular endothelial growth factor (VEGF) and their receptors. J. Cell. Sci.. 2001, roč. 114, čís. 5, s. 853–865. PMID 11181169.