C3 konvertáza
C3 konvertasa patří mezi serinové proteasy. Jako součást komplementového systému se podílí na přirozené imunitě. C3 konvertasa neboli C4bC2a.

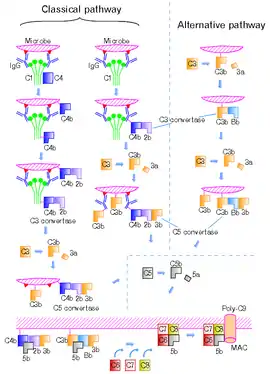
Komplementový systém může být aktivován 3 cestami – klasickou, lektinovou a alternativní. Všechny aktivační cesty vedou k enzymatickému štěpení C3 proteinu. Štěpením vzniká menší C3a a větší C3b fragment, které se uplatňují v dalších krocích komplementové kaskády. Uvolňovaný C3a působí jako chemoatraktant, a tím aktivuje zánětlivou reakci. Mezitím se C3b prostřednictvím své thiolesterové vazby kovalentně váže na mikrobiální povrch či na protilátky. C3 protein podléhá spontánnímu štěpení, tzn. i za nepřítomnosti patogena je štěpen, ale poté je jeho reaktivní vazba neutralizována molekulami vody.
Skládání C3 konvertasy
Vznik alternativní C3 konvertasy
Alternativní C3 konvertasa vzniká navázáním plazmatického faktoru B na C3b fragment, jenž asociuje s mikrobiálním povrchem. Následuje štěpení faktoru B plazmatickou serinovou proteasou faktorem D. Do plazmy je uvolněn menší fragment Ba, zatímco větší fragment Bb zůstává navázaný na C3b. Pro poskládání C3 konvertasy jsou rovněž nezbytné hořečnaté ionty.[1]
Vznik klasické a lektinové C3 konvertasy
C3 konvertasa klasické dráhy komplementu je na rozdíl od C3 konvertasy alternativní dráhy složena z jiných komplementových proteinů, a to C4b a C2a. Ty vznikají štěpením plazmatických proteinů C4 a C2 během klasické či lektinové cesty komplementu. V klasické dráze spouštění komplementu toto štěpení zprostředkovává komplex C1 (C1q, C1r, C1s) a v dráze lektinové serinové proteasy asociované s lektiny vázající manosu (MASP1, MASP2, MASP3). C4 protein je homologní k C3 proteinu a stejně jako u C3b je u fragmentu C4b štěpením odkryta vnitřní thiolesterová vazba. Protein C2 je štěpen rovněž komplexem C1 na menší (C2b) a větší fragment (C2a), který se váže na C4b. Fragmenty C4a a C2b jsou uvolněné do krve a působí jako chemoatraktanty.[1]
Funkce
- Opsonizace cizorodých částic
- Uvolňování prozánětlivých peptidů
- Formace C5 konvertasy a následná lyze buňky
Katalytická podjednotka
Katalytické podjednotky C3 konvertasy jsou v C2a a Bb podjednotkách. Jakmile jednou disociují od svých kofaktorů, už nikdy se nenavážou, aby vytvořily aktivní enzym.[2] Podjednotka Bb je tvořena dvěma doménami.[3]
Umístění na chromozomu
Geny kódující proteiny C2, C3, C4 a faktor B jsou lokalizovány na 6. chromozomu.
Regulace a stabilita
C3 konvertasy jsou nestabilní (poločas života je 10 – 20 min). Mohou disociovat spontánně či pomocí regulátorů – „faktor urychlující rozpad“ (DAF), komplementový receptor 1 (CR1), protein vázající C4b a faktor H. Proteolytické štěpení C3b (příp. C4b) brání poskládání funkční C3 konvertasy. Toto štěpení zprostředkovává v přítomnosti membránového kofaktoru (MCP, CD46), proteinu vázajícího C4b, CR1 či plazmatického glykoproteinu faktoru H komplementový faktor I. Negativní kontrolní procesy jsou nezbytně nutné pro ochranu vlastní tkáně.[4]
C3b má pro své regulátory (C3bINA, β1H, faktor B a properdin) různá vazebná místa. Vazba β1H na C3b zvyšuje vazbu C3bINA, zatímco navázaný faktor B předchází vazbě C3bINA a zároveň je kompetitivní k vazbě β1H.[5]
Pozitivní zpětná vazba
Po správném poskládání C3 konvertasa štěpí další C3 proteiny. Vznikající C3b fragmenty pak formují další C3 konvertasy. Tato pozitivní zpětná vazba je unikátním znakem pro alternativní cestu komplementu.[6] Další možností je, že konvertasa naštěpí C3 protein navázaný na již existující C3 konvertasu, tak vzniká alternativní C5 konvertasa. Pozitivní mechanismus může být regulován vazbou kontrolního proteinu - glykoproteinu β1H na C3b. β1H brání vychytávání faktoru B, usnadňuje disociační rozpad Bb, který je už navázán na C3b, a zvyšuje proteolytickou inaktivaci C3b prostřednictvím C3b inaktivátoru (C3bINA - endopeptidasa). Kyselina sialová asociovaná s membránou podporuje vysokoafinní vazbu β1H na C3b bez ovlivnění afinity faktoru B pro C3b.
Regulace amplifikační fáze alternativní cesty:
- Samovolný rozpad C3 konvertasy
- Stabilizace C3 konvertasy properdinem
- Rozpad enzymu způsobený sérovým glykoproteinem β1H
- Inaktivace C3b pomocí C3b inaktivátoru
- Ochrana C3 konvertasy před aktivací kontrolními proteiny poskytnutými povrchovými vlastnostmi buněk a ostatními aktivátory alternativní cesty
Reference
V tomto článku byl použit překlad textu z článku C3-convertase na anglické Wikipedii.
- Abbas AK, Lichtman AH, Pillai S. Cellular and Molecular Immunology.. 6th. vyd. [s.l.]: Elsevier, 2010. Dostupné online. ISBN 978-1-4160-3123-9. (anglicky)
- Kerr M. Limited proteolysis of complement components C2 and Fcator B. Biochem J. 1979, s. R615-622. (anglicky)
- Smith C, Vogel C-W, Müller-Eberhard H. MHC Class III Products: An Electron Microscopic Study of the C3 Convertases of Human Complement. J Exp Med. 1984, s. R324-329. (anglicky)
- Hourcade D, Holers V M, Atkinson J P. The regulators of complement activation (RCA) gene cluster. Adv Immunol. 1989, s. R381-416. (anglicky)
- Pangburn M, Müller-Eberhard H. Complement C3 Convertase: Cell surface restriction of β1H control and generation of restriction on neuroaminidase-treated cells. Proc Natl Acad Sci USA. 1978, s. R2416-2420. (anglicky)
- Pangburn M, Schreiber M. J Immunol. 1983, s. R1930-1935. (anglicky)