Chřipkový virus A
Chřipkové viry z čeledi Orthomyxoviridae se podle vnitřního, druhově specifického nukleoproteinového antigenu rozdělují do rodů Alphainfluenzavirus, Betainfluenzavirus, Gammainfluenzavirus, Deltainfluenzavirus.[1] Chřipkové viry typu influenza A (rod Alphainfluenzavirus s jediným druhem Influenza A virus) se vyskytují u lidí i zvířat, typy B a C pouze u lidí. Standardní nomenklatura chřipkových virů A byla navržena v roce 1971 a upravena WHO v roce 1980. Jejich označení zahrnuje typ viru, původního hostitele, geografickou oblast, číslo kmene, rok izolace a subtyp hemaglutininu (HA) a neuraminidázy (NA). Např. influenza virus typu A izolovaný u krůt v roce 1968 ve Wisconsinu, který má HA subtypu 8 a NA subtypu 4, je označován jako A/turkey/Wisconsin/1/68 (H8N4).
 H1N1 | |
| Baltimorova klasifikace virů | |
| Skupina | V (ssRNA viry s negativní polaritou) |
| Vědecká klasifikace | |
| Realm | Riboviria |
| Říše | Orthornavirae |
| Kmen | Negarnaviricota |
| Podkmen | Polyploviricotina |
| Třída | Insthoviricetes |
| Řád | Articulavirales |
| Čeleď | Orthomyxoviridae |
| Rod | Alphainfluenzavirus |
| Druh | Influenza A virus |
| Sérotypy | |
| Některá data mohou pocházet z datové položky. | |
Virologie
Morfologie
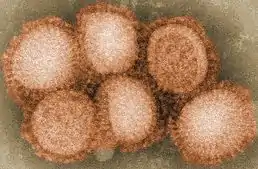
Viriony influenzových virů typu A (IVA) jsou sférické obalené částice dosahující průměrné velikosti 80-120 nm; existují i vláknité formy rozličné délky. Nukleokapsida s helikální symetrií je uzavřena ve dvouvrstevném lipidovém obalu, který obsahuje povrchové projekce délky 10-12 nm. Tyto povrchové projekce představují 2 důležité glykoproteiny kódované virem – trimer hemaglutinin (HA) a tetramer neuraminidázu (NA).
Hemaglutinin je membránový protein, obsahující hlavní povrchový antigen (H) viru. Skládá se ze dvou disulfidickou vazbou spojených řetězců (HA1 a HA2), které vznikají z prekurzoru hemaglutininu proteolytickým štěpením. Biologická aktivace HA buněčnými proteázami je nezbytným předpokladem pro infekciozitu viru, protože teprve HA2 navozuje fúzi obalu viru na plasmatickou membránu buňky. Hemaglutinin, kromě toho že odpovídá za přilnutí virionu k receptorům buněčného povrchu hostitelské buňky, vyvolává také hemaglutinaci erytrocytů a indukuje tvorbu neutralizačních a hemaglutinaci inhibujících protilátek.
Neuraminidáza obsahuje další důležitý antigen (N) viru. Je to enzym, který hydrolyticky štěpí glykosidovou vazbu mezi kyselinou sialovou (N-acetylneuraminovou) a D-galaktózou nebo D-galaktózoaminem na povrchu napadené buňky a spolupůsobí tak s HA při adsorpci a průniku viru do buňky. Uplatňuje se také při uvolňování nově vzniklých virových partikulí z buněk a při eluci viru z erytrocytů. Protilátky proti neuraminidáze pravděpodobně brání šíření infekce mezi buňkami; nemají ale neutralizační aktivitu. Na základě rozdílných antigenních vlastností HA a NA se u ptačích IVA rozlišuje 16 subtypů HA a 9 subtypů NA, které se mohou vyskytovat v různých vzájemných kombinacích.
Pod virovým obalem leží velký strukturální matrixový protein M1 obalující molekulu RNA, nukleoprotein (NP) tvořící kapsidu a 3 velké strukturální proteiny – polymerázy PB1 a PB2 a kyselý protein PA, nezbytné pro replikaci RNA a její transkripci. Virový genom představuje 8 segmentů (genů) jednovláknové RNA s negativní polaritou (-). Segmenty 1-3 a 5 kódují informace pro syntézu RNA a nukleokapsidy, segmenty 4 a 6 pro HA a NA, segment 7 pro matrixový protein M1 a segment 8 pro nestrukturální proteiny (NS1 a NS2). Segmenty RNA mohou být elektroforeticky separovány podle jejich migrační rychlosti, čehož se využívá v genetických, epizootologických i diagnostických studiích.[2]
Segment 7 od něhož se odvíjí transkript pro matrixový protein M1 je po splicingu označován jako M2. Dlouhou dobu se o jeho důležitosti vedly spory. Jelikož se jedná o hydrofobní protein, a proto k jeho sledování nebylo možné použít MRI, byla použita technika nukleárně magnetické rezonanční spektroskopie, která ukázala, že protein M2 mající tvar kanálu s póry na obou koncích musí být aktivován kyselým prostředím. Samotný proces je zahájen aminokyselinami tryptofanem a histidinem. Histidin funguje jako přenašeč protonů z buňky do nitra viru, zatímco tryptofan slouží jako vrátný, který otevírá bránu při přiblížení protonu. Jedná se o nezbytný krok k replikaci viru[3].
Přibližné složení chřipkového virionu je 0,8-1,1 % RNA, 70-75 % bílkovin, 20-24 % lipidů a 5-8 % cukrů. Lipidy jsou lokalizovány ve virové membráně většinou ve formě fosfolipidů, cukry pak ve formě glykoproteinů nebo glykolipidů.[4]
Genom
Genom je lineární, segmentovaný a enkapsidovaný nukleoproteinem NP. Jeho 8 segmentů (velikost se pohybuje od 890 do 2341 bp) kóduje 11 proteinů. Celková velikost je 13,5 kbp.
Životní cyklus
- Virus se na buňku naváže prostřednictvím HA na cukry s kyselinou sialovou.
- Buňka virus transportuje do endozómu prostřednictvím endocytózy.
- Část HA sfůzuje s membránou a uvolní tak virovou RNA (vRNA), příslušné proteiny a RNA dependentní RNA polymerázy do cytoplasmy.
- Z těchto jednotek se vytvoří komplex, který je transportován do jádra, kde RNA dependentní RNA polymeráza vytvoří komplementární pozitivní cRNA.
- Dochází k replikaci vRNA a translaci podle cRNA.
- Zvýšená koncentrace M1 indukuje export segmentů genomu z jádra pomocí NEP.
- Negativní vRNA tvořící genom budoucích virů, polymerázy a další virové proteiny utvoří virion.
- HA a NA se na povrchu buňky shluknou v bouli, virové jádro opustí buněčné jádro a vstoupí do tohoto výběžku.
- Dospělý virus vypučí z buňky obalen lipoproteinovou membránou buňky obohacenou o HA a NA, NA vyčistí novou membránu od cukrů s kyselinou sialovou.
Epidemiologie
Prevalence
Influenzavirus A způsobuje onemocnění chřipku u savců a chřipkové infekce ptáků.
K pomnožení chřipkových virů se nejčastěji používají kuřecí embrya, vzhledem k jejich vysoké citlivosti. Virusy lze adaptovat na laboratorní zvířata. Všechny kmeny viru chřipky aglutinují ptačí a některé savčí erytrocyty; jsou však rozdíly ve schopnosti aglutinovat červené krvinky určitého zvířecího druhu. K biologickým pokusům se nejčastěji používají krůťata, kachňata a kuřata.
Nebezpečí
Důležitou biologickou vlastností chřipkových virů A je schopnost vytvářet antigenní variace – antigenní posun (drift) a antigenní zvrat (shift). Antigenní posun je způsobován bodovými mutacemi částí genomu kódujících HA a NA a projevuje se malými antigenními a strukturálními změnami obou glykoproteinů. Antigenní zvrat vzniká vzájemnou výměnou segmentů nově se tvořících genomů virů (rekombinace, angl. genetic reassortment – přeuspořádání) po předchozí infekci vnímavé buňky (embrya, hostitele) dvěma nebo i více antigenně odlišnými viry. Tak dochází v přírodě ke vzniku antigenně zcela „nových“ chřipkových virů. Na vzniku antigenních variací chřipkových virů se nejčastěji z ptáků podílejí divoké kachny.
Odkazy
Literatura
- JURAJDA, Vladimír. Nemoci drůbeže a ptactva – virové infekce. 1. vyd. Brno: ES VFU Brno, 2002. 184 s. ISBN 80-7305-436-1.
- SAIF, Y.M. et al. Diseases of Poultry. 11. vyd. Ames, USA: Iowa State Press, Blackwell Publ. Comp., 2003. 1231 s. Dostupné online. ISBN 0-8138-0423-X. (anglicky)
Reference
- International Committee on Taxonomy of Viruses, Virus Taxonomy: 2019 Release. Dostupné online (anglicky)
- Swayne, D.E. a D.L. Suarez (2000): Highly pathogenic avian influenza. Rev. sci. tech. Off. int. Epiz., 19 (2) : 463-482
- Jessica Wapner, Proti chřipce, Scientific American České vydání, únor 2011, strana 16
- Choppin, P.W. a R.W. Compans. (1975). In: The Influenza Viruses and Influenza (ed. E.D. Kilbourne), pp. 15-47, Academic Press, NY
Související články
- Influenza virus B
- Influenza virus C
- Isavirus
- Togotovirus
Externí odkazy
 Obrázky, zvuky či videa k tématu chřipkový virus A na Wikimedia Commons
Obrázky, zvuky či videa k tématu chřipkový virus A na Wikimedia Commons