Mezenchymově-epitelový přechod
Mezenchymově-epitelový přechod (MET) je reverzibilní biologický proces, který zahrnuje přechod od pohyblivých, multipolárních nebo vřetenovitých mezenchymových buněk k planárním útvarům polarizovaných buněk zvaných epitel. MET je reverzní proces epitelově-mezenchymového přechodu (EMT) a ukázalo se, že se vyskytuje v ontogenezi, vznik indukovaných pluripotentních kmenových buněk ze somatických buněk jedince[1], rakovinném metastázování[2] a hojení ran[3].
Úvod
Na rozdíl od epitelových buněk, které jsou nepohyblivé a vyznačují se apikobazální polaritou s vazbou na bazální laminu, těsnými spoji, vodivými spoji, adhezními spoji a expresí markerů mezibuněčných spojů, jako je E-kadherin[4], mezenchymové buňky nevytvářejí zralé mezibuněčné spoje, mohou procházet skrz extracelulární matrix a exprimují markery, jako jsou vimentin, fibronektin, N-kadherin, Twist a Snail.[4] MET hraje zároveň zásadní roli při změnách metabolismu buněk a při upravě epigenetických modifikací. Obecně je v procesu MET postupně zvyšována exprese genů spojených s epitelovým charakterem buněk a snižována exprese genů spojených s mezenchymovým charakterem buněk.
Ontogeneze
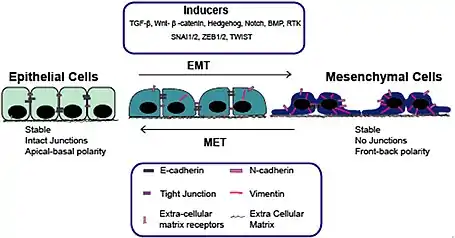
Během embryogeneze a raného vývoje se buňky přepínají tam a zpět mezi různými buněčnými fenotypy prostřednictvím MET a EMT. MET spojený s embryogenezí byl nejrozsáhleji studován na průběhu somitogeneze[5], vývoje ledvin[6] a karcinogenezi během metastazování[7]. Probíhá též během vývoje srdce[8] nebo při vývoji trávicího ústrojí[9]. MET je nezbytný proces v embryogenezi pro vytváření kohezních struktur z doposud beztvarých shluků buněk. Ačkoli mechanismus MET během vývoje různých orgánů je docela podobný, každý proces má jedinečnou signální dráhu k vyvolání změn genové exprese.
Vývoj ledvin
Jedním z nejlépe popsaných procesů využívajících MET je ontogeneze ledvin. Savčí ledvina je primárně tvořena dvěma časnými strukturami: ureterovým pupenem a nefrogenním blastémem, které tvoří sběrné kanálky a nefrony. Jak ureterový pupen roste z Wolffova vývodu, nefrogenní blastém indukuje větvení ureterového pupenu. Současně ureterový pupen indukuje kondenzaci nefrogenního blastému okolo pupenů a ten pak podstupuje MET za vzniku renálního epitelu, který nakonec tvoří nefron. V metanefronech dochází k vzájemné indukci a růstových faktorů, integrinů, buněčných adhezních molekul a protoonkogenů, jako jsou c-ret, c-ros a c-met, a k indukci MET.
Somitogeneze
Další příklad využití MET se vyskytuje během somitogeneze. Somity obratlovců, které dávají vznik kostím pánevního dna, páteři a žebrům a kosterním svalům trupu, jsou vytvářeny zráním sdružováním somitomer (další podrobnosti viz somit). Každý somit je obklopen vrstvou epitelu, který vznikl z mezenchymových buněk transformovaných pomocí MET. Na příkladu somitogeneze kuřecího embrya bylo zjištěno, že je pro správný průběh MET zapotřebí mimo jiné dvou GTPáz - Cdc42 a Rac1 a také transkripčního faktoru Paraxis.
Vývoj srdce
Během vývoje srdce probíhá několik cyklů EMT a MET. Část splanchnopleury podléhá EMT a dává vzniknout endoteliálním progenitorům, které pak tvoří endokardium prostřednictvím MET. Perikard je tvořen mezenchymálními buňkami žilní zátoky - sinus venosus, které podléhají MET. K podobným procesům dochází i při regeneraci v poškozené tkáni srdce. Poškozený perikard tedy podléhá EMT a je transformován na adipocyty nebo myofibroblasty, které mohou dále vytvářet podklad pro vznik arytmií. Následný zpětný proces v podobě MET pak vede k tvorbě vaskulogenních progenitorových buněk a epitelových progenitorů, které mohou dále diferencovat ve vaskulogenní buňky podporující angiogenezi, a tedy dochází k postupné regeneraci poškozené tkáně srdce.
V rakovině
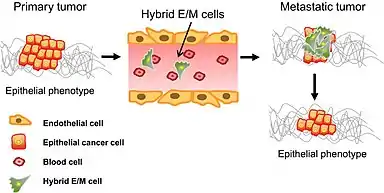
I když je známo relativně málo o tom, jakou roli hraje MET při rakovině ve srovnání s rozsáhlými studiemi o využití EMT při metastázování nádoru, předpokládá se, že se MET také podílí na vytváření a uchycení vzdálených metastáz. A to zřejmě tím, že transformovaným buňkám umožňuje znovu získat epitelové vlastnosti a schopnost integrovat se do vzdálených orgánů. Mezi těmito dvěma stavy se buňky vyskytují ve "přechodném stavu“ v tzv. částečném EMT.
V posledních letech vědci zkoumají MET jako jeden z mnoha potenciálních terapeutických cílů v prevenci vzniku metastáz.[10] Tento terapeutický přístup prevence metastáz je známý jako "diferenciační" terapie a může být použit při vývoji nových protinádorových terapeutických strategií.
Vznik iPS ze somatických buněk
Aby somatické buňky podstoupily přeprogramování na indukované pluripotentní kmenové buňky (buňky iPS), musí proběhnout řada různých buněčných procesů. Přeprogramování somatických buněk na iPS lze dosáhnout signalizací pomocí exprese Oct4, Klf4, Sox2 a c-Myc (OKSM).[11]
Geny spojené s epitelovým charakterem buněk, jako je E-kadherin / Cdh1, Cldns -3, -4, -7, -11, Occludin (Ocln), epiteliální buněčná adhezní molekula (EpCAM), a Crumbs homolog 3 (Crb3), zvýšily svou expresi dříve než byl zapnut Nanog, klíčový transkripční faktor při udržování pluripotence . Kromě toho byla během prvních 5 dnů po indukci OKSM snížena exprese genů spojených s mezenchymovým charakterem buněk, jako je Snail, Slug, Zeb -1, -2 a N-kadherin.[12] Přidání exogenního TGF-β1, který blokuje MET, významně snížilo účinnost přeprogramování iPS.[13] Všechna tato zjištění jsou v souladu s předchozími pozorováními, že embryonální kmenové buňky připomínají epitelové buňky a exprimují E-kadherin.
Nedávné studie naznačují, že signalizace pomocí Klf4 při přeprogramování buněk iPS může být konkrétně odpovědná za indukci exprese E-kadherinu vazbou na jeho promotorové oblasti a první intron CDH1, genu kódujícího E-kadherin.
Reference
V tomto článku byl použit překlad textu z článku Mesenchymal–epithelial transition na anglické Wikipedii.
- PEI, Duanqing; SHU, Xiaodong; GASSAMA-DIAGNE, Ama; THIERY, Jean Paul. Mesenchymal–epithelial transition in development and reprogramming. Nature Cell Biology. January 2019, s. 44–53. Dostupné online. ISSN 1465-7392. DOI 10.1038/s41556-018-0195-z. (anglicky).
- PASTUSHENKO, Ievgenia; BRISEBARRE, Audrey; SIFRIM, Alejandro; FIORAMONTI, Marco; REVENCO, Tatiana; BOUMAHDI, Soufiane; VAN KEYMEULEN, Alexandra. Identification of the tumour transition states occurring during EMT. Nature. April 2018, s. 463–468. Dostupné online. ISSN 0028-0836. DOI 10.1038/s41586-018-0040-3. (anglicky).
- KALLURI, Raghu. EMT: When epithelial cells decide to become mesenchymal-like cells. Journal of Clinical Investigation. 2009-06-01, s. 1417–1419. Dostupné online. ISSN 0021-9738. DOI 10.1172/JCI39675. PMID 19487817. (anglicky).
- DAS, Vishal; BHATTACHARYA, Sourya; CHIKKAPUTTAIAH, Channakeshavaiah; HAZRA, Saugata; PAL, Mintu. The basics of epithelial–mesenchymal transition (EMT): A study from a structure, dynamics, and functional perspective. Journal of Cellular Physiology. September 2019, s. 14535–14555. Dostupné online. ISSN 0021-9541. DOI 10.1002/jcp.28160. (anglicky)
- HAMIDI, Sofiane; NAKAYA, Yukiko; NAGAI, Hiroki; ALEV, Cantas; SHIBATA, Tatsuo; SHENG, Guojun. Biomechanical regulation of EMT and epithelial morphogenesis in amniote epiblast. Physical Biology. 2019-04-23, s. 041002. Dostupné online. ISSN 1478-3975. DOI 10.1088/1478-3975/ab1048. (anglicky).
- HOLMQUIST MENGELBIER, Linda; LINDELL-MUNTHER, Simon; YASUI, Hiroaki; JANSSON, Caroline; ESFANDYARI, Javanshir; KARLSSON, Jenny; LAU, Kimberly. The Iroquois homeobox proteins IRX3 and IRX5 have distinct roles in Wilms tumour development and human nephrogenesis: IRX3 and IRX5 in Wilms tumour and mammalian nephrogenesis. The Journal of Pathology. January 2019, s. 86–98. Dostupné online. DOI 10.1002/path.5171. PMID 30246301. (anglicky).
- LIAO, Tsai-Tsen; YANG, Muh-Hwa. Revisiting epithelial-mesenchymal transition in cancer metastasis: the connection between epithelial plasticity and stemness. Molecular Oncology. July 2017, s. 792–804. Dostupné online. DOI 10.1002/1878-0261.12096. PMID 28649800. (anglicky).
- NEBIGIL, Canan G.; DÉSAUBRY, Laurent. The role of GPCR signaling in cardiac Epithelial to Mesenchymal Transformation (EMT). Trends in Cardiovascular Medicine. May 2019, s. 200–204. Dostupné online. DOI 10.1016/j.tcm.2018.08.007. (anglicky).
- MU, Tianhao; XU, Liqin; ZHONG, Yu; LIU, Xinyu; ZHAO, Zhikun; HUANG, Chaoben; LAN, Xiaofeng. Characterizing the Emergence of Liver and Gallbladder from the Embryonic Endoderm through Single-Cell RNA-Seq. biorxiv.org. 2019-07-30. Dostupné online. DOI 10.1101/718775. (anglicky).
- PATTABIRAMAN, D. R.; BIERIE, B.; KOBER, K. I.; THIRU, P.; KRALL, J. A.; ZILL, C.; REINHARDT, F. Activation of PKA leads to mesenchymal-to-epithelial transition and loss of tumor-initiating ability. Science. 2016-03-04, s. aad3680–aad3680. Dostupné online. ISSN 0036-8075. DOI 10.1126/science.aad3680. PMID 26941323. (anglicky).
- Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006, s. 652–5. DOI 10.1016/j.cell.2006.07.024. PMID 16904174. (anglicky).
- Samavarchi-Tehrani P, Golipour A, David L, Sung HK, Beyer TA, Datti A, Woltjen K, Nagy A, Wrana JL. Functional genomics reveals a BMP-driven mesenchymal-to-epithelial transition in the initiation of somatic cell reprogramming. Cell Stem Cell. 2010, s. 64–77. DOI 10.1016/j.stem.2010.04.015. PMID 20621051. (anglicky).
- Li R, Liang J, Ni S, Zhou T, Qing X, Li H, He W, Chen J, Li F, Zhuang Q, Qin B, Xu J, Li W, Yang J, Gan Y, Qin D, Feng S, Song H, Yang D, Zhang B, Zeng L, Lai L, Esteban MA, Pei D. A mesenchymal-to-epithelial transition initiates and is required for the nuclear reprogramming of mouse fibroblasts. Cell Stem Cell. 2010, s. 51–63. DOI 10.1016/j.stem.2010.04.014. PMID 20621050. (anglicky).