Mechanosenzitivní iontové kanály
Mechanosenzitivní iontové kanály (zkratka MSC z angl. mechanosensitive (ion) channel) jsou typem iontových kanálů, které mění svoji konformaci a propustnost pro ionty v reakci na mechanické podráždění.
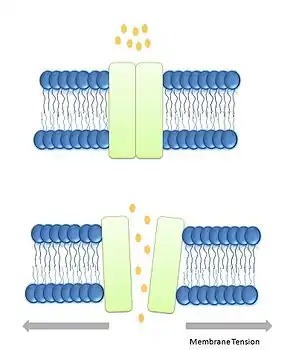
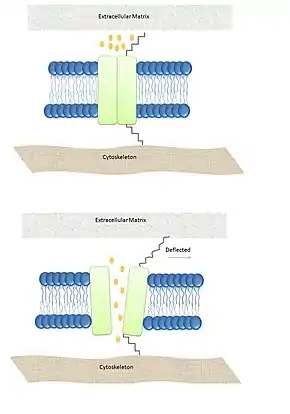
Mechanizmy otevírání kanálů
Byly popsány dva mechanizmy, kterými je mechanický signál převeden na konformační změnu bílkovinné membránové struktury.[1]
- Tzv. „intrinsic model“ vychází přímo z podráždění membrány, ve které je kanál „usazený“. Tlak cytoplazmy způsobuje natažení membrány, které vyvolá posun transmembránových domén kanálu, a tím změnu propustnosti pro ionty (viz Obr. 1). Tyto kanály využívají například bakterie pro zajištění stálého objemu cytoplazmy v hypotonickém prostředí.[2]
- Otevírání MSC na základě tzv. „trapdoor model“ je zajištěno vláknem cytoskeletu nebo mezibuněčnou hmotou připojených k některé z domén kanálu. Tímto způsobem se ovírají například kanály TRPA1, které se nachází v membráně vláskových buněk Cortiho orgánu ve vnitřním uchu. Tyto buňky fungují jako vlastní senzorický orgán, který umožňuje obratlovcům slyšet. Při procesu slyšení prochází zvukové vlny zvukovodem k bubínku a přes kůstky středního ucha jsou převedeny na vibrace kapaliny v kanálku ve vnitřním uchu. Tato kapalina prostřednictvím membrány na spodní straně kanálku (bazilární membrány) ohýbá vláskové buňky Cortiho orgánu. Kanály TRPA1 jedné buňky jsou vláknem připojeny k cytoskeletu sousední buňky. Při jejich ohybu buněk se vlákno napíná, a tím otvírá iontový kanál. Tímto kanálem začnou do buňky proudit kationty, což způsobí depolarizaci membrány a iniciaci nervového vzruchu, který putuje do sluchového centra v mozku.[3]
MSC prokaryot
Pro výzkum MSC se tradičně využívá technika patch-clamp. Ta umožňuje zjistit velikost elektrického proudu, který generuje tok iontů procházejících iontovým kanálem. Na základě velikosti tohoto proudu byly u bakterií určeny tři typy MSC - MscL (Large-Conductance Mechanosensitive Channels), MscS (Small-Conductance MSC) a MscM (Mini-Conductance MSC). Čtvrtým typem jsou kanály MscK, které jsou specifické pro ionty draslíku.[2] Nejlépe probádanou je struktura kanálu MscL. Tento kanál, vyskytující se výhradně u prokaryot, je pentamer, kde se každá podjednotka skládá ze dvou transmembránových domén. Změna vnějšího napětí se projeví otevřením póru o průměru 3 nm, kterým mohou procházet anionty a kationty. Otevření už jediného (z přibližně tisíce kanálů nacházejících se v celé membráně) vede k zaznamenatelné změně ve vnitřním prostředí buňky. Samovolnému otevření kanálu brání vysoká hodnota aktivační energie (přibližně 125 kJ/mol), nutná pro změnu jeho konformace.[4]
MSC eukaryot
Eukaryotní MSC se dělí do dvou skupin na základě iontové selektivity. První skupinou jsou kationtové kanály, mezi něž se řadí kanály Piezo, TRP a DEG/EnaC. Druhý typ je charakteristický svou specifitou pro draslíkové kationty a dvěma pórovými doménami - sem patří například kanály TREK a TRAAK. MSC se u eukaryot nachází především v membránách neuronů, ale také v orgánech, které při vykonávání své funkce musí měnit objem, jako jsou plíce, močový měchýř nebo srdce. Výše zmíněný kanál TRPA1 z vláskových buněk vnitřního ucha patří do rodiny kanálů TRP.[5]
Význam
Buňky využívají MSC k rozličným účelům. Kromě již zmíněné osmoregulace u prokaryot a sluchu obratlovců slouží také jako receptory dotyku, mechanosenzory v srdečním svalu,[6] a dokonce i jako čidla tepla a chladu v senzorických neuronech kůže.[7] Abnormální funkce MSC je spojena s některými typy srdečních arytmií a kardiomyopatií u pacientů se svalovou dystrofií,[6] hypertenzí a polycystickou chorobou ledvin.[5] Díky své lokalizaci na membráně neuronů v centrální nervové soustavě jsou také ideálním cílem celkových anestetik.[4] MSC z rodiny Piezo byly nalezeny na membránách receptorů bolesti, a existuje tedy potenciálně zajímavá možnost jejich ovlivnění pomocí nových typů analgetik.[8]
Reference
- MONSHAUSEN, G. B. a E. S. HASWELL. A force of nature: molecular mechanisms of mechanoperception in plants. Journal of Experimental Botany. 2013-11-05, vol. 64, issue 15, s. 4663-4680. DOI: 10.1093/jxb/ert204. Dostupné z: http://jxb.oxfordjournals.org/lookup/doi/10.1093/jxb/ert204
- PEROZO, Eduardo, D. Marien CORTES, Pornthep SOMPORNPISUT, Anna KLODA, Boris MARTINAC, M. J. PETRUS, A. E. DUBIN a A. PATAPOUTIAN. Open channel structure of MscL and the gating mechanism of mechanosensitive channels. Nature. 2002-8-29, vol. 418, issue 6901, s. 942-948. DOI: 10.1038/nature00992. Dostupné z: http://www.nature.com/doifinder/10.1038/nature00992
- COREY, David P., Jaime GARCÍA-AÑOVEROS, Jeffrey R. HOLT, Kelvin Y. KWAN, Shuh-Yow LIN, Melissa A. VOLLRATH, Andrea AMALFITANO, Eunice L.-M. CHEUNG, Bruce H. DERFLER, Anne DUGGAN, Gwénaëlle S. G. GÉLÉOC, Paul A. GRAY, Matthew P. HOFFMAN, Heidi L. REHM, Daniel TAMASAUSKAS a Duan-Sun ZHANG. TRPA1 is a candidate for the mechanosensitive transduction channel of vertebrate hair cells: Cardiac mechano-energetico-informatics. Nature. 2004-10-13, vol. 432, issue 7018, s. 723-730. DOI: 10.1038/nature03066. Dostupné z: http://www.nature.com/doifinder/10.1038/nature03066
- SUKHAREV a F. SACHS. Molecular force transduction by ion channels - diversity and unifying principles. Journal of Cell Science. 2012-08-28, vol. 125, issue 13, s. 3075-3083. DOI: 10.1242/jcs.092353. Dostupné z: http://jcs.biologists.org/cgi/doi/10.1242/jcs.092353
- MARTINAC, B. Mechanosensitive ion channels: molecules of mechanotransduction. Journal of Cell Science. 2004-05-15, vol. 117, issue 12. DOI: 10.1242/jcs.01232. Dostupné z: http://jcs.biologists.org/cgi/doi/10.1242/jcs.01232
- FRIEDRICH, Oliver, Soeren WAGNER, Andrew R. BATTLE, Sebastian SCHÜRMANN, Boris MARTINAC, Hideo KUSUOKA, Eduardo MARBAN, Myron L. WEISFELDT, Hideyuki SATO, Masatsugu HORI, Akira KITABATAKE, Michitoshi INOUE. Mechano-regulation of the beating heart at the cellular level – Mechanosensitive channels in normal and diseased heart: Cardiac mechano-energetico-informatics. Progress in Biophysics and Molecular Biology. 2012, vol. 110, 2-3, s. 303-317. DOI: http://dx.doi.org/10.1007/0-387-24990-7_23.
- NOËL, Jacques, Katharina ZIMMERMANN, Jérome BUSSEROLLES, Emanuel DEVAL, Abdelkrim ALLOUI, Sylvie DIOCHOT, Nicolas GUY, Marc BORSOTTO, Peter REEH, Alain ESCHALIER a Michel LAZDUNSKI. The mechano-activated K channels TRAAK and TREK-1 control both warm and cold perception. The EMBO Journal. 2009-3-12, vol. 28, issue 9, s. 1308–1318. DOI: 10.1038/emboj.2009.57. Dostupné z: http://emboj.embopress.org/cgi/doi/10.1038/emboj.2009.57
- COSTE, B., J. MATHUR, M. SCHMIDT, T. J. EARLEY, S. RANADE, M. J. PETRUS, A. E. DUBIN a A. PATAPOUTIAN. Piezo1 and Piezo2 Are Essential Components of Distinct Mechanically Activated Cation Channels. Science. 2010-09-30, vol. 330, issue 6000, s. 55-60. DOI: 10.1126/science.1193270. Dostupné z: http://www.sciencemag.org/cgi/doi/10.1126/science.1193270